Example of a well-written lab report
Return to Laboratory report Instruction main page
Example of a poorly written lab report
(single-spaced to conserve paper; yours should be double-spaced to leave room for comments)
Ontogenetic Color Change and Mating Cues
in Largus californicus (Hemiptera: Largidae)
Carey Booth Box 123
Biology 102 2 February 1995
Lab instructor: Ned Knight
Lab day: Friday
Abstract
Ontogenetic color change at sexual maturation can be useful in identifying an appropriate mate for some organisms. Largus californicus individuals undergo two ontogenetic color changes. First instars are bright red, second through fifth instars are shiny blue-black, and adults are black with orange markings. Adult male mating behavior suggested that the change in color from fifth instars to adults might enable males to discriminate between nymphs and adults. Males mount adults and persist if they have mounted a female and quickly release if they have mounted another male. Males were never observed to mount nymphs. Female color patterns were altered and male's copulatory attempts were timed to determine if color pattern was used by males in mating decisions. The null hypothesis that dorsal color pattern does not significantly affect male mating behavior could not be rejected, therefore the significance of the color change from nymph to adult must be sought elsewhere.
Introduction
Ontogenetic color change at the time of sexual maturation has been shown to be advantageous to fish (Fricke 1980), reptiles (Werner, 1978), and birds (Lyon and Montgomerie, 1986). In general, dull-colored juveniles avoid predation risk and harassment by breeding males, and sexually mature individuals use bright colors to advertise their readiness to mate (Booth, 1990a). In insects, mating cues are often chemical rather than visual (Jacobson, 1972), but there are some exceptions. In diurnal Lepidoptera, adult color pattern plays an important role in the initial phase of mating behavior (Graham et al., 1980). In holometabolous insects, such as Lepidoptera, maturation is associated with dramatic morphological changes, therefore distinguishing between larvae and adults for mating attempts is not difficult. The recognition of maturity is more difficult in hemimetabolous insects where late instars may be similar to adults in size and shape. The possibility that ontogenetic color change in some Hemiptera may have evolved as an indication of maturation has not been investigated experimentally.
The mating behavior of male Largus californicus suggests that males may be using visual cues, perhaps in addition to pheromonal cues, to distinguish between fifth instars and adults for mating attempts. Fifth instars are shiny blue-black and almost adult-sized. Adults (both males and females) are black with orange borders around the thickened portion of the hemelytra and pronotum (Booth, 1990b). Although males were never observed to mount nymphs, they do mount other adults, and persist if they have mounted a female or release within a few seconds if they have mounted another male. Their distinctive courtship behavior allows an observer to identify immediately the initiation of a mating event. This consists of the male orienting towards the female when he is approximately 1 cm away, rapidly waving his antennae, leaping onto the female's back, and agitatedly grabbing the female with his legs. As their genitals do not immediately join, it is possible to separate a pair before they actually mate.
These bugs do not fly and are easily handled and painted without significantly disrupting their normal behavior. Experiments were designed to determine if males use color cues in their mating decision and if their behavior could explain the significance of the ontogenetic color change from fifth instars to adults. The null hypothesis that dorsal color pattern does not significantly affect male mating behavior was tested.
Materials and Methods
The experiment was performed outdoors at the Main Campus Reserve at the University of California, Santa Barbara on January 31, 1988. Bugs were collected from the Reserve on the morning of the testing day. Tests were performed when the bugs are normally active (1030 to 1430 hours) and control tests were interspersed between experimental tests so that time of day, temperature, cloudiness, and other environmental variables would be approximately the same between experiment and control.
An acrylic black paint and clear finish were used in each treatment. The first treatment was black paint and clear finish on the ventral surface of the female to control for the smell of the paints without altering the black and orange pattern on the dorsum. The second treatment was clear finish on the dorsum to control for covering the dorsal surface, which may reduce any scent emitted or otherwise affect the female's behavior. The third treatment was black paint on the dorsum to mimic the color of the fifth instars. The three treatments will be referred to as normal, clear, and black for brevity.
One female was used for all three treatments to hold other aspects (size, shape, scent) of the female's attractiveness constant. The order of presentation of the three treatments was necessarily the same for all males, as the one female in each experiment could only have black paint added after the normal and clear treatments. This design allows for a repeated measures analysis of variance as each male is tested with the same female under three different paint conditions.
After each painting, the female was placed in a clear plastic 9 x 7 x 3 cm box. Males were held separately in labeled plastic petri dishes. Each male was introduced one at a time into the box at the point farthest from the female. He was removed when he mounted the female or after an arbitrarily chosen time of 270 seconds had elapsed, whichever came first. The time to mount or 270 seconds (no-mount) was recorded. The pair was separated before their genitalia joined so no actual mating occurred. After all 15 males were tested, the female was painted for the next treatment and the males were tested in the same order.
To control for the possibility of males tiring by the second or third trial, a similar number of different males were tested three times each with one untreated female; i.e. no changes were made to the female between trials. Trials were alternated between experimental and control males throughout the day of testing. A total of two females (one experimental and one control) and 29 males (15 experimental and 14 control) were used.
Statistical analyses were performed using the StatView program on a Macintosh microcomputer. One-way, repeated measures ANOVAs were used to test for differences in males' time to mount among the three treatments and among the three control trials.
Results
No significant differences were found in males' time to mount among the three treatments or among the three control trials based on a repeated measures ANOVA (Table 1). Males mounted black painted females as readily as females with the typical black and orange pattern.
There was a slight, but not significant, increase in male's mean time to mount for the black treatment as compared to the normal and clear treatments (Figure 1). The 95% confidence intervals were also larger for the black treatment. The first control trial had a slightly larger, but not significantly different, male's mean time to mount as compared to the second and third trials (Figure 2). The male's mean times to mount were lower for the three control trials than for the three experimental treatments.
Because the maximal time males were allowed to stay in the box without mounting the female was chosen arbitrarily, the one case where a male did not mount the female within the allotted 270 seconds could have biased the results (Table 1). After excluding the mount time for the male that failed to mount, the results did not differ qualitatively from the above: no significant differences were found.
Discussion
By using one female for all three color treatments, any non-color aspects of the female's attractiveness were held constant. As the null hypothesis (that males' time to mount is not significantly affected by color of the female) was not rejected, males evidently used those other traits in seeking a mate. The male behavior of mounting other adults (male or female) and not nymphs may indicate that there are pheromonal differences between nymphs and adults but not between adult males and females. Males release other males rapidly once contact has been made, so chemical cues transferred by touch or other close range signals (such as sound) may be used to distinguish males from females. There are slight shape differences between nymphs and adults (nymphs are more spherical) that could possibly be used by males in mating decisions. Other experiments are necessary to determine the nature of the communication between adults and between adults and nymphs.
Among hemipterans, several species use pheromones as mating cues. Males of the southern green stink bug (Nezara viridula) release a pheromone that attracts females, males, late-stage nymphs, and a parasitoid (Aldrich et al., 1987). Females of Dysdercus cingulatus and Pyrrhocoris apterus also produce substances attractive to males (Osmani and Naidu, 1967; Zdarek, 1970). As these last two species are in the same superfamily (Pyrrhocoroidea) as L. californicus, it is possible that L. californicus females also produce a pheromone that is attractive to males. However, several species in the family Largidae, including L. cinctus (a close relative of L. californicus), have minimal development of the metathoracic scent gland evaporative area (Schaefer, 1972), so their use of pheromonal communication may be limited. The use of pheromones does not rule out the possibility that visual cues may also be important.
References
Aldrich, J. R., J. E. Oliver, W. R. Lusby, J. P. Kochansky and J. A. Lockwood. 1987. Pheromone strains of the cosmopolitan pest, Nezara viridula (Heteroptera: Pentatomidae). J. Exp. Zool. 244: 171-175.
Booth, C. L. 1990a. Evolutionary significance of ontogenetic colour change in animals. Biol. J. Linn. Soc. 40: 125-163.
Booth, C. L. 1990b. Biology of Largus californicus (Hemiptera: Largidae). Southwestern Naturalist 35: 15-22.
Fricke, H. W. 1980. Juvenile-adult colour patterns and coexistence in the territorial coral reef fish Pomacanthus imperator. Mar. Ecol. 1: 133-141.
Graham, S. M., W. B. Watt and L. F. Gall. 1980. Metabolic resource allocation vs. mating attractiveness: Adaptive pressures on the "alba" polymorphism of Colias butterflies. Proc. Natl. Acad. Sci. 77: 3615-3619.
Jacobson, M. 1972. Insect sex pheromones. Academic Press, New York.
Lyon, B. E. and R. D. Montgomerie. 1986. Delayed plumage maturation in passerine birds: reliable signaling by subordinate males? Evolution 40: 605-615.
Osmani, Z. and M. B. Naidu. 1967. Evidence of sex attractant in female Dysdercus cingulatus Fabr. Indian J. Exp. Biol. 5: 51.
Schaefer, C. W. 1972. Degree of metathoracic scent-gland development in the trichophorous Heteroptera (Hemiptera). Ann. Entomol. Soc. Am. 65: 810-821.
Werner, D. I. 1978. On the biology of Tropidurus delanonis, Baur (Iguanidae). Z. Tierpsychol. 47: 337-395.
Zdarek, J. 1970. Mating behaviour in the bug, Pyrrhocoris apterus L. (Heteroptera): ontogeny and its environmental control. Behaviour 37: 253-268.
Table 1. Repeated measures ANOVA on males' time to mount female (in seconds).
|
Treatment or Trial |
number males |
mean timea ± SEMb |
number no-mounts |
ANOVA Between Treatments
|
||||||||||||||||
|
|
|
|
1.42 2,28 0.26 | ||||||||||||||||
|
|
|
|
1.34 2, 26 0.28 |
a Males' copulatory attempts were timed from point of entry into female container.
b SEM = standard error of the mean
c F = F statistic
d df = degrees of freedom for numerator, denominator
e P = probability value
f Each Largus californicus male was tested with the same female under three different paint conditions. Black and clear paint on the dorsum = Normal treatment. Clear paint on the dorsum = Clear treatment. Black paint on the dorsum = Black treatment to mimic the color of the fifth instar larva.
g Different males were tested three times each with one untreated female = Control trials 1-3.
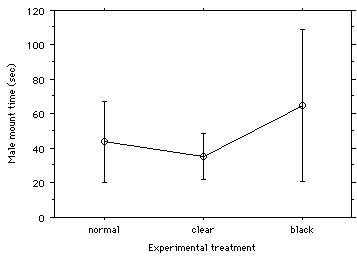
Figure 1: Mean male mount time (sec) with 95% confidence intervals under three experimental conditions. Each Largus californicus male was tested for time to mount (attempting copulation) with the same female under three different paint conditions. Normal = Black and clear paint on the dorsum to control for odor of paints. Clear = Clear paint on the dorsum to control for covering the dorsal surface. Black = Black paint on the dorsum to mimic the color of the fifth instar larva.
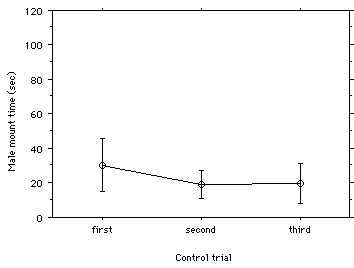
Figure 2: Mean male mount time (sec) with 95% confidence intervals for three control trials. Different Largus californicus males were tested for time to mount (attempting copulation) three times each with one untreated female to control for order of presentation in the experimental treatments.