Eggshell Removal in Shorebirds
Biology 342 Fall 06
By Molly & Jennifer
| Home | Mechanism | Ontogeny | Phylogeny | Adaptive Value | References | 2006 websites |
Phylogeny
Parents of birds with nidiculous young, or chicks that remain in the nest after hatching, almost universally remove eggshells after chicks have hatched. Birds with nidifugous young, or young that leave the nest soon after hatching, generally do not bother to remove the eggs. Both behavior patterns achieve the same goal, however—getting the young far away from the egg shells.Shorebirds are a curious exception to this pattern. Many have precocial young that leave the nest soon after hatching, but some of these shorebirds will still remove the shells of hatched eggs from the nest. It is possible that this trait is “left over” from their evolutionary history. However, research on members of the Recurvirostridae family has shown that egg removal is still an advantage in precocial birds because predators will often attack nests before the birds leave. Precocial birds that are not in the shorebird family, such as ducks, do not remove eggshells after hatching, but tend to remove broken eggshells during incubation. The tendency to remove eggshells from the nest after incubation seems to depend largely on how developmentally advanced the young are at the time of hatching. Duck chicks can leave the nest almost immediately, while precocial shorebirds will wait 1-2 days to leave the nest. Researchers have investigated this trend in nidifugous shorebirds; see Evolution for hypotheses on the adaptive advantages of this behavior.
Kittiwakes of the family Laridae (also Larini) do not remove their eggs at all. This is thought to be a result of the fact that they breed in cliffs, where egg shells are unlikely to make their nests any more conspicuous. This is especially interesting because they are in the same family as the black headed gulls and other gulls that nearly universally display this egg removal behavior. Since the rest of the Laridae family exhibits the eggshell removing trait, it is likely that the Kittiwakes independently lost this trait, since assuming that all of the other species independently gained the trait would be a violation of the parsimony principle.
Members in all three major branches of modern shorebirds have the egg removing behavior. This means that the trait was likely possessed by their common ancestor. Since the trait has been maintained in many modern shorebirds, it has likely contributed to the fitness of these birds over evolutionary time.
Evaluating the Shorebird Phylogenies
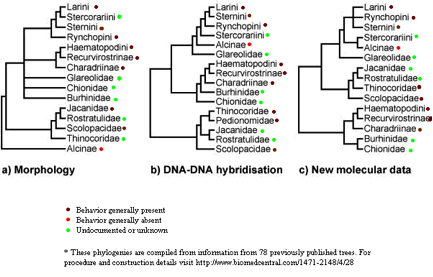
The Alicidae family (made up of auks and murres), the fourth branch on most traditional morphological shorebird phylogenies is not included as a separated branch when DNA differences and other molecular analyses are used to create the phylogeny. Morphological phylogenies take into account unique structures and behaviors of each family and attempt to group them based on those characteristics. The danger with morphological grouping is that it is difficult to determine to what extent a different trait reflects distant speciation, and it is often hard to distinguish homologous and homoplastic traits when making these groupings. Recently, molecular data has been favored in making phylogenies because the number of different base pairs between two species can be used as an index of how related these species are and how long ago they diverged. In closely related species, such as the shorebirds, DNA analysis is favored since it experiences a higher rate of mutation than other molecular data (such as RNA and proteins). Since DNA has many noncoding regions, mutations occur in these areas more quickly, and this can be used along with other data to analyze how long ago the species diverged.
A central but highly challenged method in constructing morphological phylogenies is called the parsimony principle. This principle states that when constructing phylogenies, if there are multiple plausible ways to relate the organisms to each other, one should use the construction that requires the fewest independent gains or losses of a trait to have occurred. This principle makes sense theoretically since mutations, and as a result, morphological changes are the product of chance errors in transcribing the DNA sequence. Since it is less likely in a purely probabilistic sense for the same genetic sequence to mutate twice, the parsimony principle is a good rule of thumb in constructing phylogenies. However, as we can see from the DNA-DNA hybridization model, the parsimony principle does not always reflect genetic similarity.
Let’s take the eggshell removal trait as an example (excluding the undocumented species in our analysis). If we were simply using the parsimony principle and morphological traits as our guideline, we probably would come up with a phylogeny similar to the one seen in a. If eggshell removal behavior developed right after the Alicidae family speciated, then we would not need to assume any independent gains or losses, and as a result would have constructed the “best” phylogeny according to the parsimony principle. But the DNA-DNA hybridization phylogeny shows us that the Alicidae family is closely related to other families that exhibit the eggshell removal behavior. As it turns out, the species in the Alicidae family live on cliffs, much like the kittiwakes (the lone Laridae species that does not exhibit eggshell removing behavior). Since the Alicidae is a species closely related to eggshell removing shorebirds, it likely independently lost the behavior relatively recently. This loss tells us something about the environment in which the Alicidae live and the value of eggshell removal in general. As we have seen in the kittiwakes, cliff-dwelling birds are subject to little interspecific predation and need not camouflage their nests from predators. Eggshell removal in this species would leave the chicks unprotected with no resulting anti-predation advantage. Therefore, a strong selectional pressure against eggshell removal could develop, resulting in the independent loss of such a trait.
Each of the above phylogenies gives us different information about how the birds are related to each other. By considering this information together, we can learn more about the relatedness of one species to another, and by evaluating independent gains and losses among closely related animals we can hypothesize about the unique selectional pressures that group experienced in its evolutionary history.
Much family-specific shorebird information was found on http://bna.birds.cornell.edu/BNA/