Kleptoparasitism
Biology 342 Fall 2012
Erin Appleby and Ryan Streur
Mechanism
Mechanism is one of Tinbergen’s four questions which pertains to the causes of a behavior at a moment in time. Little to no research has been done on mechanism related to kleptoparasitism in seabirds, but we can understand this behavior by looking at the mechanisms that contribute to the more general food search behavior: 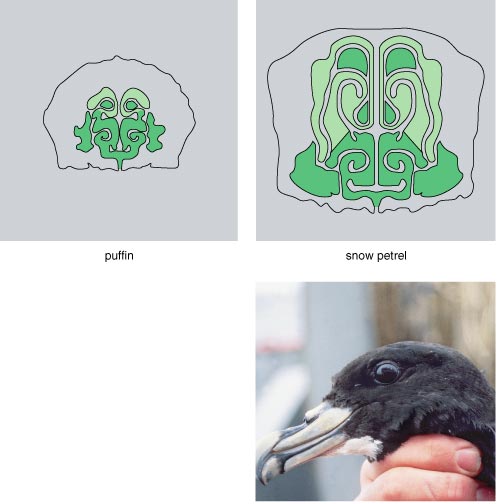
- location of hosts by a highly-developed sense of smell
- brain structures that enable higher cognitive processes to innovate foraging techniques
- hormonal fluctuations that influence foraging behavior
The Sense of Smell in Tube-nosed Seabirds
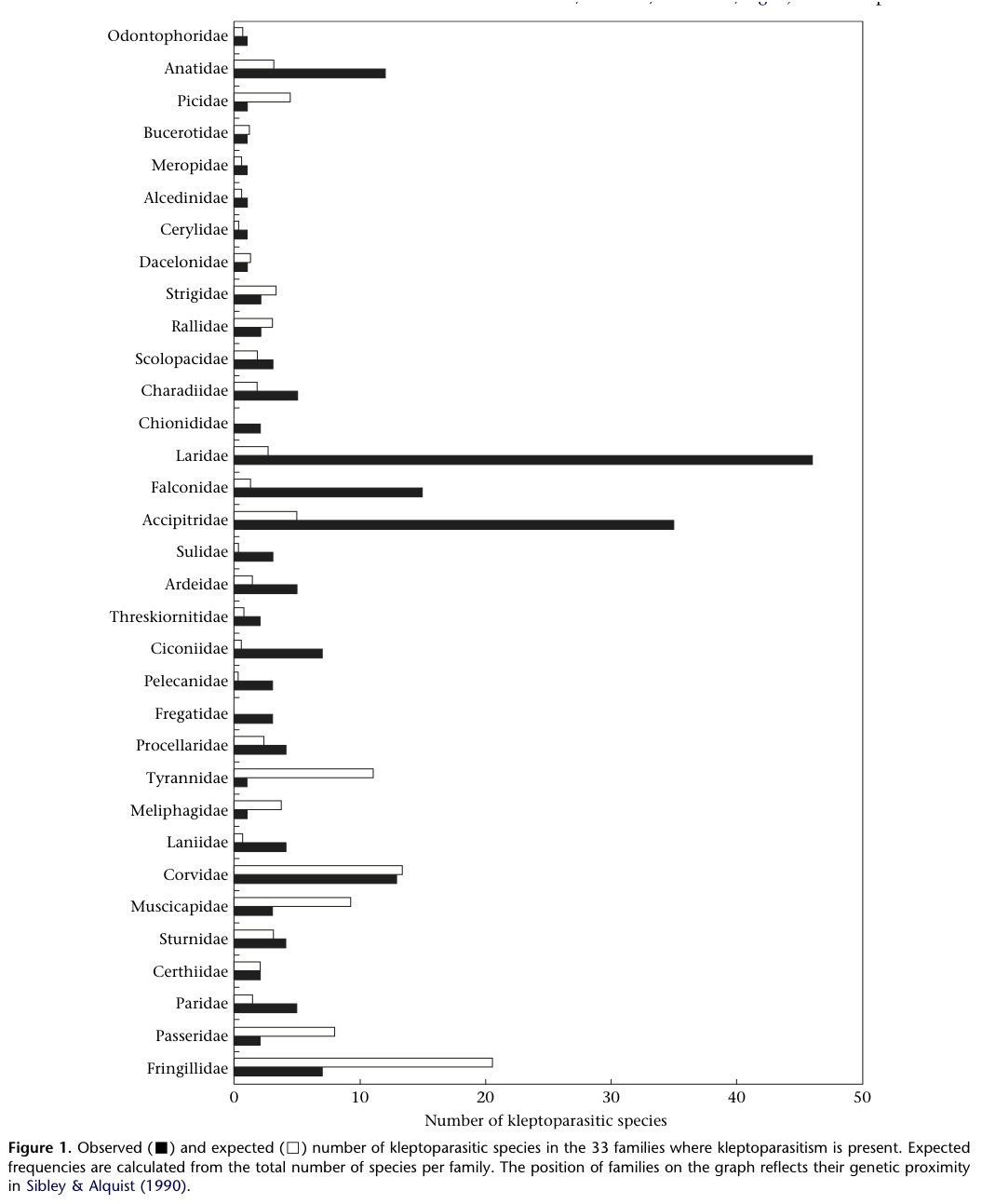
Procellariiform seabirds include petrels, shearwaters, and albatrosses--species known to be kleptoparasitic (See PHYLOGENY or Morrand-Ferron bar graph). Alternatively referred to as “tube-nosed” for their characteristic tubular nostrils on the upper bill, this order of birds has the largest olfactory bulbs (smelling-organs) of all birds (Nevitt, 2008). In addition, the architecture of their brains suggest a highly developed olfactory sense (Britannica “procellariiform”). To the right, the brain structure devoted to the sense of smell is compared between a puffin and a snow petrel (a procellariiform).
{kind=link}
When a tube-nosed bird is foraging for food over vast areas of ocean, it locates “hot spots” where prey is likely to be found by responding to the scent of dimethyl sulfide (DMS). DMS is a byproduct of the metabolic processes of phytoplankton, and large quantities of it is released when they are being fed on by zooplankton. Phytoplankton and zooplankton are microscopic organisms abundant in the ocean, which support most marine food chains. It has been shown that many foraging marine animals, including tube-nosed birds are able to detect the scent of these compounds. A high concentration of zooplankton attract predators such as squid, a tasty treat for seabirds. The plumes of the odor (shown as odor feature below) indicate to foraging animals that predators of zooplankton such as squid are likely to be found in that patch (Nevitt, 2008). So the same olfactory mechanism that allows a seabird to detect a prey-rich patch of ocean also facilitates the detection of a patch that will likely be host-rich as well.
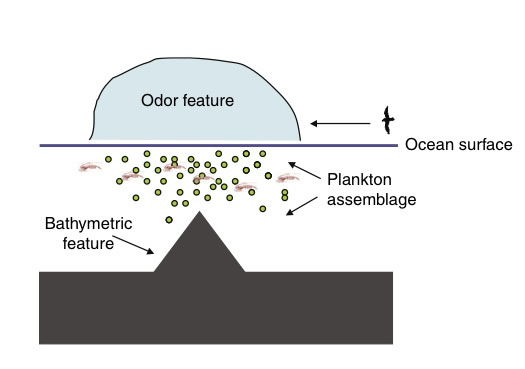 “Predators tend to be messy eaters, and procellariiform birds are adapted to pay attention to who is eating whom” (Nevitt 2008).
“Predators tend to be messy eaters, and procellariiform birds are adapted to pay attention to who is eating whom” (Nevitt 2008).
Once the “hot spot” is found, tube-nosed birds switch to small-scale foraging, (spans tens to hundreds of square kilometers) and begin to rely on both sight and smell to locate and capture prey (Nevitt 2008). When it comes to small-scale foraging, birds are not necessarily attracted to the scent of their prey (such as krill or squid) but to DMS and other odor indicators of elevated feeding activity.
However, albatrosses and giant petrels do respond to the scent of crushed krill and the activity of other birds and marine predators. These tend to be larger more aggressive birds as well. One would expect this extra attention to “who is eating whom” would give these birds important information to enhance kleptoparasitic success. Further research in this scope may shed light on the following question:
Do seabirds employ the same sensory mechanisms used to scan and hunt for prey as they do when scanning for hosts (Hamilton 2002)?
Studies that address this question can use the below model to examine psychological mechanisms in the same manner we might examine a physical one, such as wing morphology. The below model is used to support the hypothesis of convergent evolution of intelligence in apes and corvids, a taxonomic family of birds. The well-developed brain structure of the scrub-jay is linked to its impressive cognitive ability to hide food stores, anticipate the hiding places of other birds, and steal their food stores (More on this in PHYLOGENY). Launching from this foundation, further research can be done to learn about cognitive mechanisms in a seabird species to expand our knowledge on other cognitively-demanding behaviors, such as kleptoparasitism, on a mechanistic level.
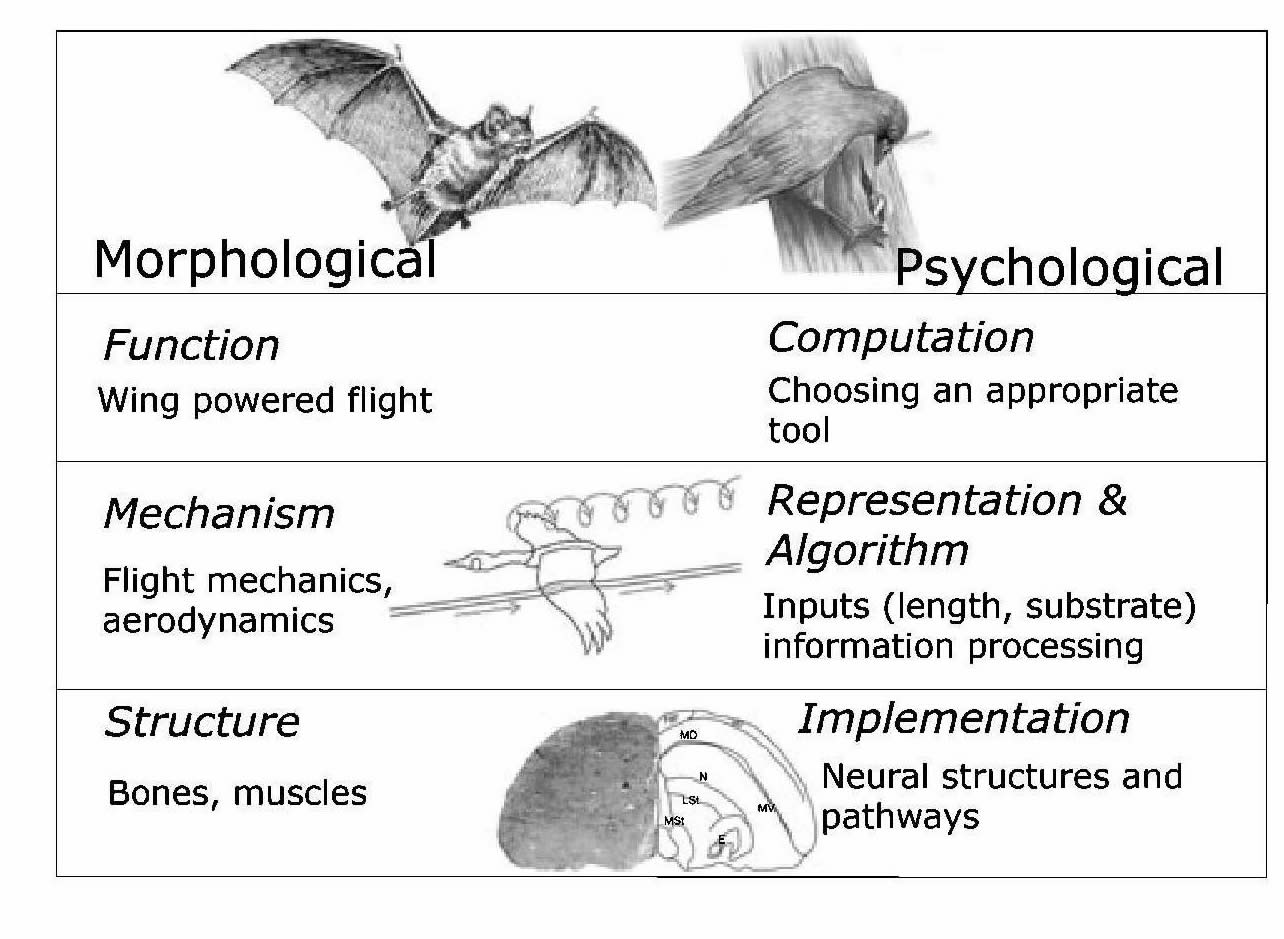
Above: A model that organizes psychological mechanism into three levels of complexity analogous to the physical mechanism (Marr 1982).
Hormones Influence Foraging
Foraging behavior is often influenced by food scarcity and breeding season. Breeding impacts foraging since the offspring are another mouth to forage for in an ocean with unpredictable food sources. In addition, when it comes to breeding, it is quality over quantity for seabirds (small brood size, slow growth rate). Corticosterone, a stress hormone, has been suggested to influence foraging behavior in the common tern (Sterna hirudo) in studies on breeding ecology and parental care. Short-term elevated levels of glucocorticoids, another stress indicator, during chick-rearing may be linked to greater foraging effort to support their young (Reichert 2012). Chick-rearing male black-legged kittiwakes (Rissa tridactyla) responded to elevated corticosterone levels with an increased time spent foraging/flying, but only in males in good body condition (Angelier 2007).