Bio 332 - Vascular Plant Diversity:
EXAM 1 KEY
The following answers are given as guidelines to the kinds of information I was looking for. Other correct answers may have been possible for some questions, so you should check with me if you have specific questions.
1. Ernst Mayr (1992, Am. J. Bot. 79: 222-238) provided both a philosophical defense of the Biological Species concept and an empirical analysis based on applying this definition to the local flora in Concord, NH.
Do you conclude that Mayr has effectively made the case for botanical use of the BSC?
If you say “yes”, indicate what criticisms directed at the BSC were dealt with effectively by Mayr’s results. If you say “no”, indicate the problems you see with Mayr's choice of data collection methodology and/or with the reasoning he used to reach his conclusion
various answers/justification possible.
YES – Mayr examined the kinds of taxa for which the BSC has genuine difficulties (asexual reproduction, hybridization, taxa with various ploidies that make interbreeding difficult to judge) and found that these reflected the minority of species in the Concord Flora.
NO – Mayr applied a definition that invokes reproductive isolation and interbreeding without any experimental support from performing empirical tests of interbreeding. He used morphology in many cases to infer the degree of interbreeding. He tackled the species question in a setting without concern for spatial variation (the extent to which interbreeding occurs for geographically distinct populations of a species) and without concern for temporal variation. His study also focused on a flora that is extremely well-studied, but it’s not clear if this would lead to a biased conclusion about species definitions (i.e., does study of a flora minimize the occurrence of asexual reproduction and hybridization?). Given his methods, a well-studied flora does seem to have been a requirement for the study he conducted.
2. In their report on sympatric speciation in Howea palms, Savolainen et al. (2006; Nature 441: 210-213) begin their article with a quote about allopatric speciation (attributed to Turelli et al., 2001):
“...the conceptual rationale is simply that, given enough time, speciation is an inevitable consequence of populations evolving in allopatry”.
Give a full description of the temporal and geographic context and the underlying evolutionary genetics that might make allopatric speciation inevitable. Include in your description what role could be played by natural selection, and if this role is required
Populations in allopatry and without connection through gene flow are expected to diverge genetically over time. This will happen because of the inevitable effects of mutation, coupled with genetic drift and natural selection. If epistasis is a general feature of genomes and governs the way genes interact with each other, we can expect the genetic divergence over time will lead to the accumulation of Dobzhansky-Muller-Bateson incompatibilities. These interacting loci will contribute to post-zygotic reproductive isolation whenever the two populations interbreed and the epistatic effects of the DMB gene loci become expressed in the hybrid offspring. In addition, natural selection may lead to the divergence of traits that directly affect reproduction and will also therefore contribute to post-zygotic or pre-zygotic reproductive isolation. The consequence of genetic divergence, over time in allopatry, will therefore inevitably lead to the inability of the populations to interbreed successfully, which results in speciation
3. McNeilly (1968; Heredity 23: 99-108) investigated the joint effects of gene flow and natural selection in a study of copper tolerance for the grass, Agrostis tenuis, near and on a copper mine in Wales. The figure below shows copper tolerance measured in adult plants (filled bars) and their offspring (open bars) collected from locations on and off the mine.
A third transect (the green line) has been added to the map, beginning in the meadow off the mine and continues onto the mine. The transition between the meadow and mine occurs midway along this transect (indicated by the green arrow). As before, adults and offspring are collected from positions along this transect (indicated by the six asterisks on the figure at the bottom).
For this new set of six sampling points, indicate on the graph above your expectation for the copper tolerance that would be observed for both the adults (solid bars) and their offspring (open bars) measured at each sample point.
Also provide a brief description of the roles that selection and gene flow have played in producing the results you have drawn
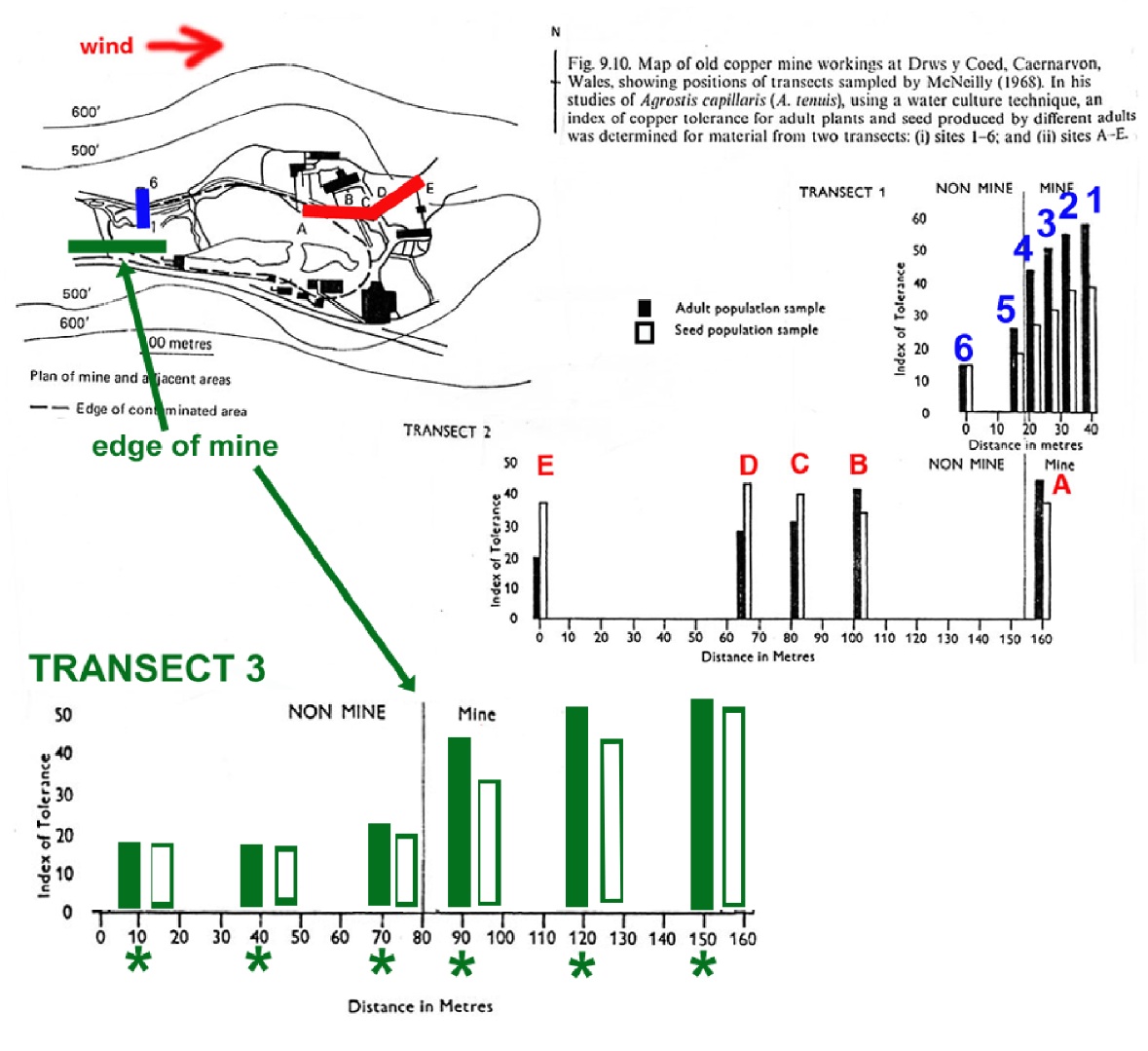
Natural selection off the mine favors low copper tolerance (because of the competitive advantage for non-tolerant plants). On the mine, selection favors copper tolerance in the plants. The other factor important in this transect is the influence of gene flow, which is predominantly occurring from off the mine towards the mine. The offspring of parents at any of the six sampling sites will reflect the effects of gene flow, which in most cases are carrying "non-tolerance" alleles, so the offspring will be generally less tolerant than the parents.
The first two sampling sites (off the mine) show adults that are non-tolerant (therefore locally adapted) and offspring that reflect gene flow from other non-tolerant individuals. The third site near the mine border may show increased tolerance in the adults as a result of selection to match increased levels of copper that may be in the soil near the mine's edge, but the offspring will have less tolerance due to gene flow from the non-tolerant individuals upwind and off the mine. For the three sampling sites on the mine, the tolerance shown in the adults will be high, while the offspring will be less tolerant due to gene flow from off the mine. The difference between parents and offspring will be greatest near the mine boundary where gene flow from off the mine is most effective. Further into the mine, there will be less gene flow from the non-tolerant plants off the mine which will result in both parents and offspring showing greater and more similar adaptation through copper tolerance to the mine's soils
4. Les Gottleib (1973, Am. J. Bot. 60: 545-553) observed that Stephanomeria malheurensis differs from its presumed ancestor (S. exigua subsp. coronaria) by at least a half-dozen traits, including the absence of a seed dormancy that is normally broken in S. e. subsp. coronaria only when conditions are favorable for seed germination and seedling survival.
Describe an evolutionary mechanism that could account for the evolution of a non-adaptive trait (e.g., the loss of seed dormancy) during the formation of this new species. Also indicate if a role for this evolutionary mechanism helps to identify the geographic mode (i.e., sympatric vs. allopatric) by which S. malheurensis originated from S. e. subsp. coronaria.
Traits that are non-adaptive (or even maladaptive) will not go to fixation as a result of selection alone. Genetic drift is one mechanism that can lead to fixation of deleterious alleles. (Though less likely - it is also possible that indirect effects of the trait – perhaps due to linkage with other, favored traits – might contribute to the increase in the non-adaptive trait). Drift is potentially consistent with either speciation mode if the trait under consideration is not responsible on its own for reproductive isolation.
Sympatric divergence creating new species is unlikely to happen by drift alone, so evidence of drift does not do much to support sympatric speciation. However, the sympatric speciation could have occurred due to selection favoring a trait like self-fertility; once the new species became reproductively isolated due to the selfing breeding system (favored by selection), its population size may have remained small enough for drift to result in the fixation of maladaptive traits.
Allopatric speciation might also involve the action of drift, leading to fixation of deleterious traits. This might be especially likely at the edge of the original species’ range, where population sizes may crash in some years due to inhospitable environmental conditions. The bottleneck would lead to the fixation of the maladaptive allele, and the absence of the original species during this process would have limited the opportunity for gene flow to prevent the loss of the dormancy allele
5. Orchid species are world-renowned for the intricate morphological features of their flowers. In natural communities, reproductive barriers exist that effectively prevent the formation of hybrids even when different orchid species are observed to flower at the same time. In cultivation, by contrast, orchid fanciers have been able to freely interbreed the various species, resulting in the production of spectacular (and often fertile) hybrid orchids.
Provide an evolutionary hypothesis for how speciation might proceed in orchids. Be certain to clearly specify i) the geographic circumstances (i.e. "mode" of speciation) and, ii) the evolutionary process by which the isolating barrier originates
Mechanical and Ethological are the most likely current RI barriers (the question suggests they must be pre-mating barriers, and temporal and geographic are ruled out since they are in the same communities and flowering at the same time).
In allopatric populations, selection by the local pollinator species has resulted in local adaptation to that pollinator and consequently ethological and mechanical features of the flower that will aid in pollination by the pollinators specific to that habitat. When these orchid populations have subsequently come back into sympatry (secondary contact), they are now isolated species by virtue of the mechanical/ethological changes in their floral traits that results from local adaptation.
(NOTE: the evolution of mechanical/ethological barriers in sympatry is less likely because of the difficulty in splitting a single, randomly mating population into two isolated populations through disruptive selection that focuses on adaptation to different pollinators).
6. Ellstrand et al. (1996; PNAS 93: 5090-5093) surveyed the floras of five regions from the Northern Hemisphere and determined the number of naturally occurring, hybrid taxa (including allopolyploid species) that were present. In each flora, they found the hybrid taxa were concentrated in a few families and genera, and they observed that hybridizing taxa were characterized by a number of similar traits.
In four of the regions (Scandinavia, the British Isles, and the Great Plains and the Intermountain West of North America), the percent of Families with hybrids ranged from 21 to 34%. The fifth flora in the report was Hawaii, where the percent of Families with hybrids was observed to be lower (16%).
Using your knowledge of speciation and hybridization, provide an explanation as to why there are fewer hybrids reported in Hawaii.
The authors of the article did not provide a specific answer to this question. Patterns such as this can be explained by factors that contribute to hybrid formation and/or to hybrid persistence once formed. Hawaii might differ because it is more tropical (closer to the equator), because of the geographic structure of the islands, because it is a relatively young flora, because it was not as disturbed by recent climate change, or because it is isolated relative to the continental floras.
(Formation)
Geography – If most of the reproductive isolation is provided by pre-mating barriers, there may be few opportunities for hybrids to form. In an area that is a collection of small islands, most species diversity occurs between islands (allopatry) rather than within islands. The pre-mating geographic barriers give related species few opportunities to hybridize.
Isolation – Hawaii may have a lower incidence of outcrossing taxa (a trait which the authors found to be associated with the incidence of hybridization). It could be expected that fewer outcrossing taxa are likely to make it to Hawaii since the islands are most likely to be colonized by self-fertilizing taxa because of their ability to establish a population with few (or one) plant.
Age – Hawaii has a young geologic age for its flora -- hybrid accumulation may be a consequence of the age of a region, with Hawaii's recent origin providing less time for hybrid accumulation. Alternately, if hybridization is characteristic of recent disturbance, the northern floras were heavily disturbed by the ice age that ended ~12,000 years ago, so they may show higher hybridization that results from new opportunities for gene flow for taxa that were separated during the ice age.
(Persistence)
Tropical - Hybrids are generally more frequent as you move away from the equator, and Hawaii is closer to the equator than the other floras. It may be that stronger biotic selection occurs in tropical and sub-tropical areas like Hawaii -- strong biotic selection may provide less forgiving conditions for the persistence of hybrid taxa if their fitness suffers. Alternately, the harsher abiotic conditions of the more northern floras might provide conditions that are more favorable to hybrids when they have improved fitness over their parents.
7. In the Sierra Nevada Mountains of California, two species of columbine are found that differ in aspects of their ecology and reproductive biology, but which hybridize in areas where they come into sympatry (Fig. c). Aquilegia formosa is red-flowered and holds its flowers in a pendent (facing downward) fashion (Fig. a). This species tends to be found at lower elevations, it possess petal "spurs" containing nectar that are relatively short, and it is principally pollinated by hummingbirds. A. pubescens, by contrast, possesses flowers that are a paler, whitish color and are held in an erect, upright fashion (Fig. b). The species is found at higher elevations, possesses longer nectar spurs, and is pollinated primarily by hawk moths.
Hodges and Arnold (1994, PNAS 91: 2493-2496) collected data across an elevational gradient where the two species were found to be hybridizing. They measured the morphology of flowers at the different sites, and also measured genetic variation using genetic markers coded for by nuclear DNA. These markers had alternate forms (band presence/absence) that were diagnostic of the two species, and the markers were assumed to represent "neutral" variation that is not affected by natural selection.
The changes in the frequency of the floral morphology and genetic marker traits across the zone of hybridization are shown below: In the case of the morphology (figure on the left), all traits showed a very sharp transition from flowers that looked like A. formosa to those that looked like A. pubescens. For the molecular genetic markers (on the right), the transition was much more gradual, with the hybrid zone having plants with mixtures of the alleles from the two parent species.
i. What evolutionary factor(s) is/are contributing to the pattern observed for the molecular markers?
If the markers are truly neutral, then the primary influence shaping their distribution is gene flow. Genetic drift might also be affecting divergence between the elevation extremes if either species involves population sizes that are small.
ii. What evolutionary factor(s) is/are contributing to the pattern observed for the floral morphology traits?
If gene flow is affecting the neutral markers, then it is also having some effect for the floral traits. The pattern of divergence shows a much steeper transition for morphology, suggesting that another factor is at work -- most likely this would be natural selection, which appears to be favoring the A. formosa floral traits at the intermediate elevation.
iii. Describe a research program (e.g. data you might collect or experiments you might perform) that would help you to predict the long-term, evolutionary fate of this hybrid zone. Be specific in your answer when indicating how the experiment or data collected will distinguish among possible evolutionary outcomes of the current hybridization.
Many possible answers here. Hybridization and introgression can have a variety of outcomes. In most cases, the fate will depend on how the fitness of hybrid offspring will compare to the fitness of either parental species. Hybrid fitness could be examined in the greenhouse to examine fitness effects that are not environmentally dependent, but it would most likely be helpful to examine the fate of the hybrids in a variety of sites (parental and the intermediate elevation) to see whether their fitness is higher than or lower than the parent's. Predicted outcomes could include persistence of the hybrids if their fitness is higher in a limited range, or at the other extreme, reinforcement and the evolution of additional pre-zygotic barriers if the hybrids are less fit.