Hive Behavior in Honey Bees
James Holland and Theresa Steele
Biology 342
Hive Dynamics
Evolution, Sex, and Caste
Honey bee colonies typically consist of about 20 to 40 thousand workers, zero to few thousand males (drones), depending on the time of year, and a single queen, the mother of the colony (Page et al). Because only the queen can reproduce, natural selection acts upon both individuals and colonies. Each trait or gene within the population of the hive must be adaptive on one or both of these levels (Tarpy et al 2004). Hive construction and maintenance as well as establishment of a new hive (swarming) are traits overwhelming favoring colony cooperation, whereas this altruistic model breaks down when the colony must select a new queen (Tarpy et al 2004, Wray et al 2011).
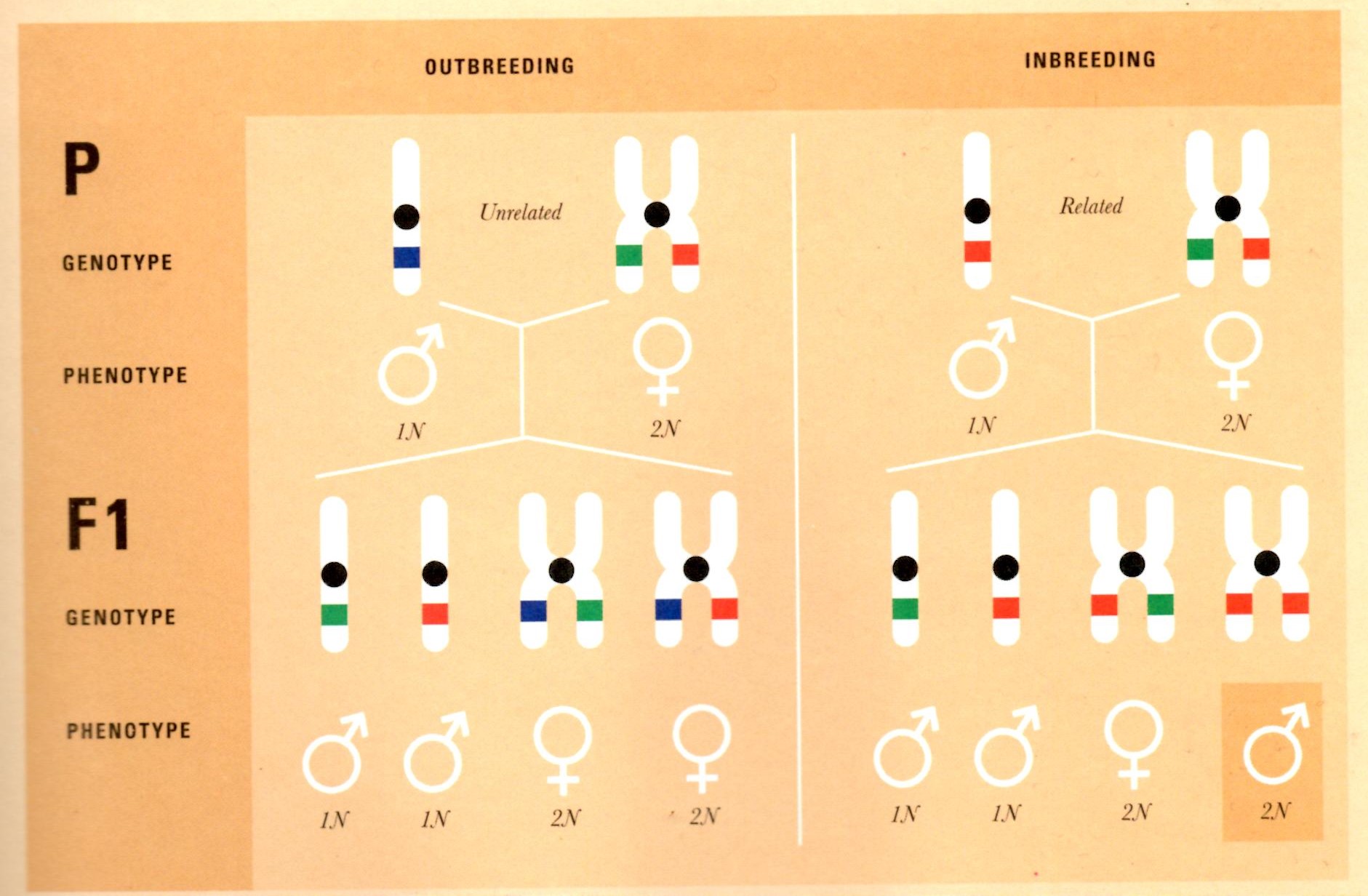
A depiction of haplodiploid reproduction, demonstrating how easily this system can produce very inbred females without sufficient outbreeding. From Wilson-Rich et al. 2014.
Honey bee sex is determined by a single locus haplodiploidy system in which the male drones contain single chromatids in their cells and the female workers and queens contain chromosomes in their cells. Unfertilized eggs laid by the queen will mature into haploid drones with half the queen’s somatic genome whereas eggs that have been fertilized by drones will mature into diploid females with a mix of the queen and the drone’s genes. Because the workers are descended from a diploid queen and a haploid drone, they have a genome that is 50% similar to one parent and 100% similar to the other. Consequently workers have genomes that are 75% similar to each other. While this promotes cooperation and altruistic like behaviors, it leaves the hive's gene pool vulnerable to accumulating deleterious recessive alleles. This is warded against by ensuring the queen usually mates with a foreign drone (outbreeding) rather than its own child (inbreeding) (Wilson-Rich 35).
The more-than-siblings degree of relatedness between worker bees is exceptional in the animal kingdom and was probably a significant driving force behind the original evolution of eusociality. While not all eusocial animals employ haplodiploidy, the magnified cooperation resultant from the very average relatedness between the workers may allow for honey bees and other haplodiploid eusocial animals (i.e. hymenoptera) to achieve such massive colony sizes (Gadagkar et al 1990). Haplodiploidy also causes all inheritable attributes of parental care to be optimized for the female only diploid genome. Genes and alleles relevant to brood rearing evolve to function in a diploid system, whereas male genes, which do not need to optimize for long life or any non mating traits, evolve to function in a haploid system. Genetic drift and time to allele fixation and loss is also significantly reduced in the lineage for this reason (Reeve et al 1993).
When a queen has died and QMP levels in the colony decrease, workers with small ovaries initiate the rearing of new queens (from eggs laid by the old queen), while more reproductively mature workers --who are generally less responsive to QMP signalling -- will activate their ovaries and lay haploid eggs, engaging in far less queen rearing behavior (Galbraith et al 2015).
Swarming
Development of swarm consensus in a single honey bee hive. Figure from Seeley et al. 2006
At a certain point in the development of a hive a collective decision is made to fission off into a mother and daughter hive through a process known as swarming. Swarming usually occurs in late spring when the colony has an excess of workers and has become overcrowded. In order for a new colony to form, a new queen must be reared from the brood and taken along with workers, honey, and a new generation of larvae to a new nest site. Usually the majority of the workers flee the nest with the queen and begin a highly organized process of scouting, transportation, and selection. Two variables must be optimized during this process: reaching the new hive before the swarm runs out of honey and the worker population begins to dwindle and die, as well as ensuring the new nest site can support the colony in the years to come. In order to accomplish the risky and computationally mammoth task of selecting a spot from all potential spots within range of the mother nest, honey bees employ a bottom up system of communication similar to the means by which they communicate foraging information to each other.

A Honey bee swarm. Image from Queen Anne View.
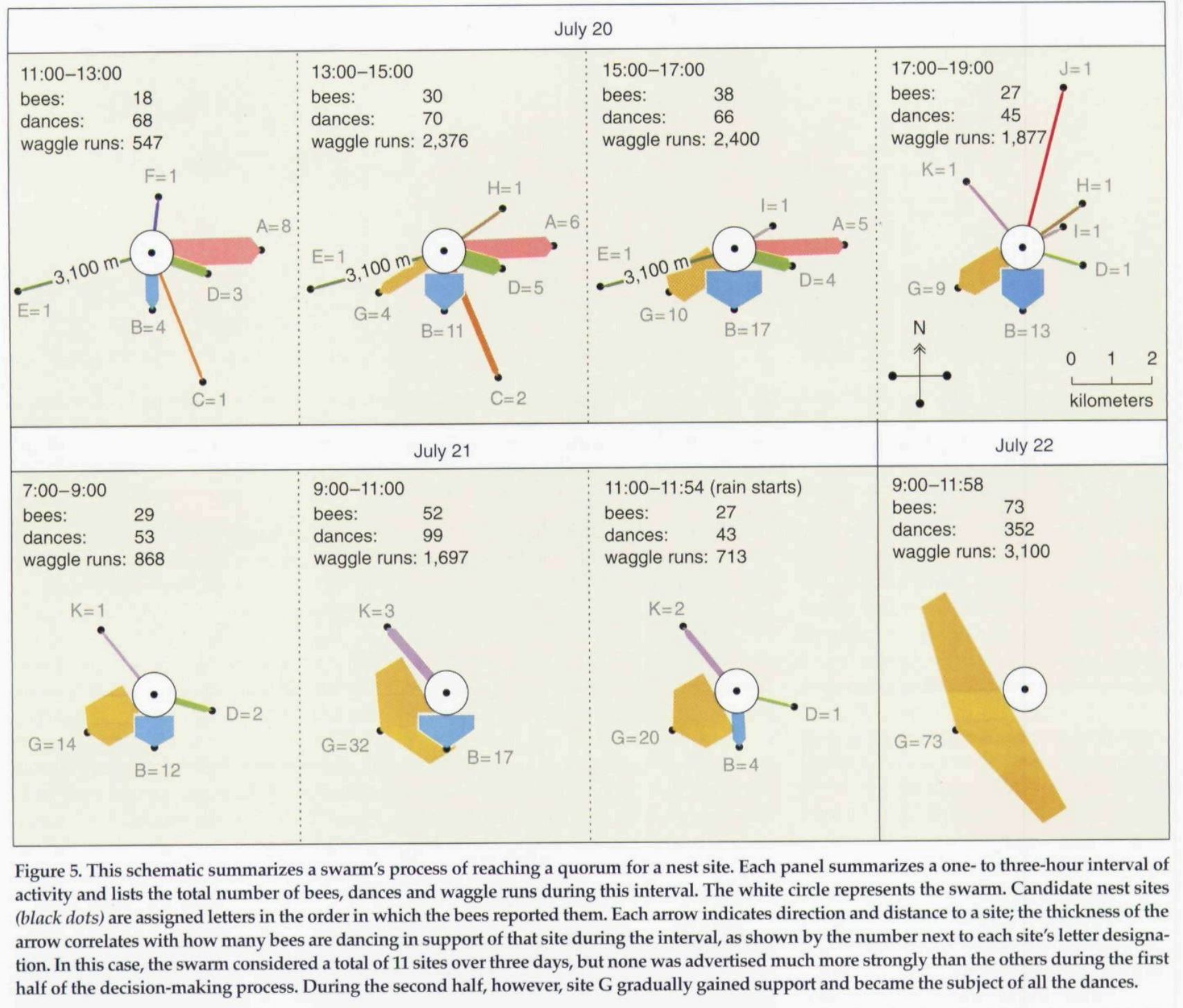
As soon as the swarm have broken off from the mother hive they rest at an interim site usually not far from their old home. At this point, or potentially as much as 3 days before the swarm breaks off from the mother nest, a series of scouts are sent out to perform reconnaissance missions by inspecting potential nest sites. The scout inspects the locale itself under variable environmental conditions (e.g. times of day, temperatures) and also communicates its findings to other scouts via physical contact and by exchanging pheromones. As the scouts return to the interim nest they begin to dance against their swarm mates to indicate the location and relevant features of their potential nest site.However, different scouts will communicate about different sites, sending conflicting messages to the swarm thereby causing their neighbors to dance in disparate fashions, as the non-scout workers mirror the dances of the scouts. Consequently, at first many different dances are performed but eventually less favorable sites cease to be danced about until at last a single site remains in contention. Once this consensus is reached the swarm flies to the new site (Winston 72).