Cuttlefish Camouflage
Biology 342 Fall 2014
Emily Merfeld & Nicolette Tapia
The Mechanism of Camouflage
-
What cues cause the cuttlefish to change color, texture, or pattern?
Visual, rather than tactile, cues are responsible for changes in color, texture, and pattern (Allen et al., 2009). These signals originate from highly light-sensitive and perceptive eyes (Messenger, 1981). The cuttlefish’s appearance changes depending on environmental light quality (e.g, frequency and amplitude), light direction (e.g., above or below), light contrast, and spatial scale (Marshall and Messenger, 1996; Mathger et al., 2006; Barbosa et al., 2007). The eyes of the cephalopods are sensitive to any orientation of polarized light through retinal irregularities (Tasaki and Karita, 1966) or specific eye movements (Shashar and Cronin, 1996). The detection of polarized light allows for a private communication channel between cuttlefish (Shashar et al., 1996).
While their perception of light contrast and quality is extremely detailed, cuttlefish are actually colorblind (Brown and Brown, 1958; Bellingham et al., 1998; Mathger et al., 2006). The mechanisms of camouflage in low-contrast, colorful environments remain to be elucidated (Mathger et al., 2005).
-
What are the cells responsible for skin color and texture changes?
The dual action of chromatophores and structural reflector cells reflect light in a large variety of ways, giving the cuttlefish a large repertoire of optical effects (Williams, 1909; Schafer, 1937; Cloney and Brocco, 1983; Messenger, 2001). The chromatophore is a small, pigmented organ surrounded by radial muscles. The color of chromatophores is controlled by rapid contraction and relaxation (see Figure 1 and video below) of radial muscles (Florey, 1969), and the proportion of expanded chromatophores determines the color of the cuttlefish. When chromatophores expand to create a dark spot, the surrounding skin pales. This is associated with decreased firing in the nerve that stimulates the surrounding area, suggesting an inhibitory relationship between the spot and the surrounding area. Artificially innervating the nerve that correlates to the dark region does not cause paling, suggesting that this inhibitory relationship is centrally controlled (Miyan et al., 1986).
See chromatophores in action. Video from Deravi et al., Royal Society Interface journal supplements.
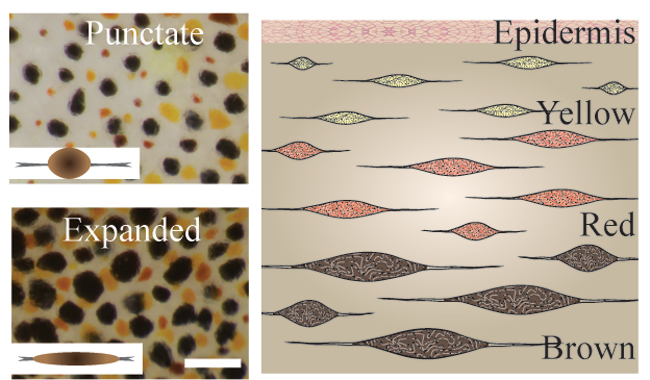
Figure 1. Punctated and expanded chromatophores, controlled by the contraction and relaxation of radial muscles. Images courtesy of Lydia Mathger.
- How are these signals interpreted, and how fast does a physical change occur?
Color change is regulated by neural, rather than hormonal signals (Boycott, 1953, 1961). Specifically, visual information is interpreted in the optic lobes, peduncle lobes, lateral basal lobes, and eventually the chromatophore lobes (Figure 2; Messenger, 2001). Radial muscles are innervated directly by the brain and alter chromatophore size in less than one second (Hill and Solandt, 1935), providing the cuttlefish with rapid camouflage that may adapt quickly to new environments.
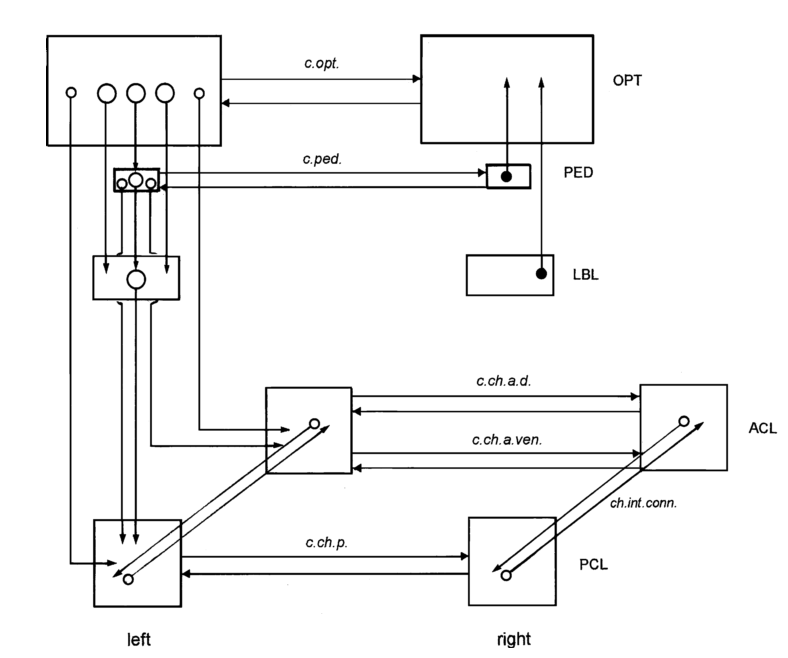
Figure 2. Simplified schematic of neural control of body patterning in cephalopods. The optic lobe, peduncle lobe, lateral basal lobe, and anterior and posterior chromatophore lobes are of particular importance, as represented by their size. Image from Messenger et al., 2001 and based on data from Young (1971), Camm (1986), and Messenger.
- How are skin patterns formed, and what do they look like?
Selective expression of chromatophores allows for pattern formations, such as stripes and spots, to match the environment (Hanlon, 1982; Mathger and Hanlon, 2007). Patterns are typically grouped into three categories: uniform, mottle, and disruptive (Figure 3). Uniform patterns are characterized by their low contrast variation, mottle patterns by their homogenous, grainy contrast variation, and disruptive patterns by their large, heterogeneous blotches (Barbosa et al., 2007). A variety of environmental variables, including size, contrast, and configuration of stimuli is factored into the expression of each pattern (Barbosa et al., 2007; Chiao et al., 2010; Hanlon and Messenger, 1988; Shohet et al., 2006).

Figure 3. The three major camouflage patterns: uniform, mottle, and disruptive. Image courtesy of Roger Hanlon.
- How are textures formed on the surface of the skin?
Papillae on the skin’s surface, which take a variety of sizes, shapes, and colorations, account for the range of skin texture from smooth to spiky (Hanlon and Messenger, 1998; Hanlon, 2007). Hydrostatic muscular control allows the relatively elastic dermis to stimulate muscle fibers, which in turn erect the papillae. When they contract, dermal erector muscles push the papillae up from the skin’s surface and to a point (Allen et al., 2013). Thus the papillae takes shape by either lying smoothly on the skin’s surface or extending away from it (Figure 4), depending on environmental conditions such as substrate (Figure 5).
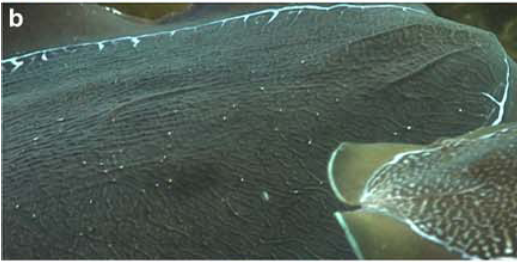
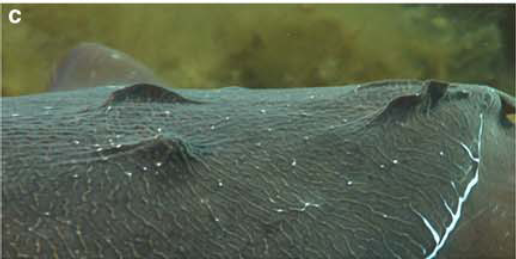

Figure 4. From left to right: Smooth skin, partially expressed papillae, and strongly expressed papillae Image from Allen et al., 2009.
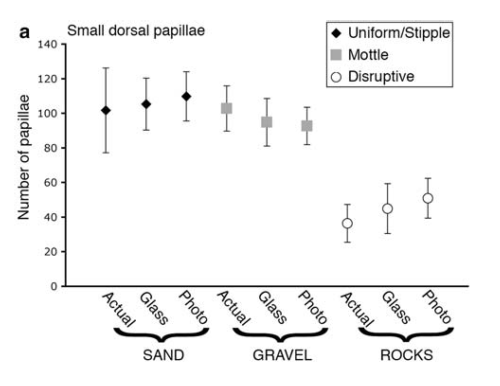
Figure 5. Small dorsal papillae expressed for each substrate type. Not shown are various expression levels for major lateral mantle papillae and major lateral eye papillae. Image from Allen et al., 2009.