Complex Cognition in Corvidae
Giving "bird brain" a whole new meaning...
| Home | Phylogeny | Mechanism | References |
If men had wings and bore black feathers, few of them would be clever enough to be crows.
—Rev. Henry Ward Beecher
Mechanism
The Physiology Behind Corvid Intelligence, in Brief, and How it Works, at Length
Corvid brains are considerably larger than would be predicted by body size, with ravens having the largest relative brain size of any bird (Emery & Clayton 2004a). In fact, the brains of crows and ravens compare in relative size to those of great apes. This measure of 'relative size' controls for body weight, and is termed the encephalization quotient (EQ), following Jerison (1973). EQ appears to correlate with cognitive ability, and in crows and chimpanzees this measure is identical. This comparison, however, is flawed, because all vertebrate brains consist of fore-, hind-, and midbrains, and these divisions are reflective of different functions. The midbrain and hindbrain are responsible for processing sensory information and organizing movement, and there is relatively little variation among species. The forebrain is the seat of conscious activity, and is implicated in sensation, learning, memory, and mood. This part of the brain varies greatly among species, but in this index too, corvids top the charts. Corvid forebrains are significantly larger than in other birds (excepting some parrots), especially in areas that are functionally analogous to the mammalian prefrontal cortex (Rehkamper et al. 1991). In mammalian brains, the prefrontal cortex is the locus of planning, decision-making, personality expression, and social behavior (Dalley et al. 2004), and is significantly enlarged in great apes credited with complex cognition. In avian brains, the analogous areas (the nidopallium and mesopallium) are significantly enlarged in corvids (Rehkamper et al. 1991). Interestingly, the mammalian neocortex is also derived from the pallium (Jarvis & Consortium 2005). This may reflect an increase in corvid intelligence similar to that of the great apes.
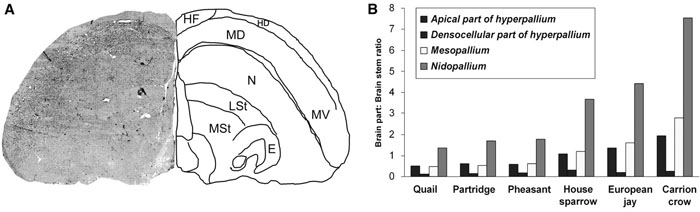
Despite their analogous functions, however, the structure of these mammalian and avian seats of cognition are very different. In mammals, the top two-thirds of the brain —the neocortex—consists of six layers of nerve cells (laminar arrangement). Bird brains, however, are formed from clusters of neurons gathered together into separate structures called nuclei (nuclear arrangement). For over a hundred years, it has been thought that brains lacking a laminar cortex must necessarily be dedicated to unconscious and instinctual behaviors, but in recent research Corvidae has been challenging this line of thought, suggesting that there is more than one way to wire an intelligent brain. Pepperberg (1999) provides a useful computer analogy when comparing mammalian and avian brains; mammalian brains are like IBM-PCs, whereas avian brains are like Apple Macintoshes; the wiring and processing are different, but the resulting output (i.e. behavior) is similar.
Intelligence Illustrated:
Following Emery (2004b, see homepage), complex cognition can be thought to comprise a "tool-kit" of tell-tale cognitive abilities: understanding cause-and-effect relationships, mentally simulating things that have already occurred or that might occur, and acting on information flexibly by generalizing rules and making inferences instead of rote learning. There is considerable overlap of these categories in each particular behavior considered.
Causal Reasoning and Behavioral Innovation
Although many animals, particularly birds and primates, use tools (Beck 1980), it is yet unclear whether any completely appreciate how tools function. Using tools to achieve an end implies causation, but not necessarily an understanding of it. However, If any non-human species could be said to understand the implied causality of tool use, it is the New Caledonian crow, Corvus moneduloides. Holzhaider et al. (2010) puts it bluntly: "New Caledonian crows have the most complex tool-manufacturing abilities among nonhuman animals, including primates." At natural foraging sites in the South Pacific, these crows whittle twigs into hooks and fashion leaves into barbed probes that they then use to extract bugs from crevices, carrying these tools with them while foraging (Hunt 1996). In the laboratory, the profundity of these observations is being sounded out. Betty, a crow in the lab, modifies a material she would never encounter in the wild, a straight, thin gauge metal wire, into a more useful crook-like hooked structure in order to lift a bucket (a cup with a handle) containing food out of a vertical tube (Weir et al. 2002). When presented with wires of different lengths and widths, she correctly selects the right tool for the job. To perform such an action, Betty would have needed to understand the problem of removing the bucket from the well to access the food, the inadequacies of a straight tool, and the flexible properties of the wire itself. This suggests several levels of causal understanding, and in fact, with the possible exception
of rooks (Bird & Emery 2009), they appear to be the only nonhuman species known to have solved problems
requiring tool use through causal reasoning (Holzhaider et al. 2010).
The rook (Corvus frugilegus), a member of the corvid family that does not appear to use tools in the wild, is also able to solve complex problems in the lab. In an experiment by Chris Bird and Nathan Emery, four captive rooks were presented with the problem of raising the water level in a container so that a worm floating on the surface moved into beak's-reach. Not only did all four solve this problem, but they learned that larger stones are most effective, and that sawdust cannot be manipulated in the same manner as water. Subject birds had no experience of this or any similar phenomena prior to testing, and never demonstrated any such behavior in captivity. The experiment demonstrates the rook's flexible ability to use tools (Bird & Emery 2009, see video)
How Insightful!
Insight is well-defined as "the sudden production of a new adaptive response not arrived at by trial behaviour or as the solution of a problem by the sudden adaptive reorganization of experience" (Thorpe 1964). In two experiments, Bernd Heinrich shows that common ravens are capable of such insight.
In the first, individual ravens were presented with a novel situation: meat, suspended from the branch of a tree using string (note that in the the string portion of the experiment above, the crows learned to do this task in training sessions, not spontaneously) In order to access the meat, the solution required "perching above the string and the food, reaching down, pulling up a loop of string, setting the looped-up string onto the perch, stepping onto the string, releasing the string with the bill while simultaneously applying pressure with the foot onto it, then reaching down again to repeat the cycle six to eight times in that precise order" (Heinrich 1995). Though his subjects in the experiment were hand-reared and possibly exceptional, some successfully completed this sequence on their first trial, with no practice with, or exposure to, the correct sequence of intermediary steps. Not only could they handle this novelty, but other novelties that were introduced, such as crossing the strings, changing the string's color, or attaching one string to food and the other to a stone, still resulted in the meat being pulled up on the first attempt. Heinrich concludes that insight is the most parsimonious explanation-- that is, the findings cannot adequately be accounted for by innate genetic programing (since this situation is completely novel; the birds had never even seen string before), learning processes (because no trail-an-error was required; no conditioning), or random chance (see video above).
In another, he compared peck marks left on suet, a common winter food for corvids, by Blue Jays, American Crows, and Common Ravens. He found that although most ravens fed like the jays and crows, at least one individual raven made distinct grooves, aligning dozens of consecutive pecks in order to cut off transportable chunks. This is suggestive of planning on the part of the raven, in order to cache this previously uncachable food source (Heinrich 1999).
Meta-tool use (using one tool to get or use another) is a tall order in the world of animal cognition; its emergence in the hominid tool-use record (in the form of stone-knapping) is an important landmark for anthropologists. In a 2009 study at Oxford University, New Caledonian crows were required to use a tool to retrieve a second tool, then use the second tool to retrieve a third, and finally use the third tool to access the food. Four out of the seven birds tested successfully used tools in a sequence from their first trial without pre-training on any particular step of the sequence, though success did increase with experience. The researchers conclude that this sequential tool use "reveals a competence beyond that observed in any other species..."(Wimpenny et al. 2009), but they were reluctant to attribute this to analogical reasoning. They cited simpler processes, such as conditional reinforcement, that would be sufficient. A follow-up study conducted at the University of Aukland the next year Taylor et al. (2010) addressed whether or not conditional reinforcement, cited in the above, could provide a complete explanation. They conducted a similar sequential tool-use experiment with only two tools, but instead of laying the first tool in front of the tubes containing other tools, they suspended it from a string. This was to serve as negative reinforcement in the training sessions, because when the bird tried accessing the food with this stick, it was unable to do so. This too-short stick would be associated with ineffective tool use, and therefore labeled an unattractive object. Crows with previous experience of each discrete behavior component (pulling up string to get short a short stick, using a short stick to access a long stick, and using a long stick to access food) linked them into a novel sequence to solve the problem on the first trial. Crows with experience of only using string and tools to access food (no pre-exposure to the middle step) also successfully solved the problem. The researchers conclude that "this innovative use of established behaviours in novel contexts was not based on resurgence, chaining and conditional reinforcement. Instead, the performance was consistent with the transfer of an abstract, causal rule: 'out-of-reach objects can be accessed using a tool'."
Self-awareness and Awareness of Others
In apes, self-directed behavior in response to a mirror has been taken as evidence of self-recognition (Gallup 1970; Povinelii et al. 1997), but it was recently found that the magpie was capable of the same (Prior et al. 2008). Magpie necks were marked with a yellow dot, and behaviors were scored with exposure to a mirror. Birds always exhibited mark directed behavior (attempt to reach it with beak, touching it with the foot) and this behavior ceased as soon as the mark was removed. Mark-directed behaviors were absent when birds were tested without a mirror. This is the first evidence of self-directed behavior in a non-mammal.
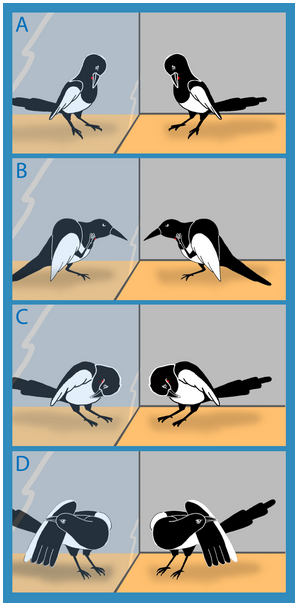

(A) attempt to reach mark with beak; (B) touching mark area with foot; (C) touching breast region outside mark area; (D) touching other parts of the body. Behaviors (A) and (B) were considered "mark-directed"
while (C) and (D) were considered "self-directed" but not directed at the mark.
Episodic Memory and Theory of Mind
Most corvids cache food for future consumption. Clark's nutcrackers can cache up to 30,000 pine seeds in a broad area, and are able to remember them up to six months later (Balda & Kamil 1992). While an amazing feat in it's own right, these birds are only demonstrating memory of the "what" and "where" of the caching event. Western scrub-jays (Aphelocoma californica), however, have been shown to be able to differentiate between caches of perishable and non-perishable food by keeping track of their relative rates of decay (Clayton & Dickinson 1998). This is the first evidence of an animal exhibiting something like episodic memory. Not only did they remember what foods they had cached and where they had cached them, they differentially recovered them depending on the perishability of the item and how long ago they had cached it. If enough time had passed that a perishable cache would have spoiled, the birds don't bother to dig up this cache and only retrieve the non-perishables and the ones that haven't yet perished. Scrub-jays apparently remember the "what-where-and-when" of specific caching events in the past. The authors of this study argue that such performance fits the behavioral bill for episodic memory, a capacity not yet demonstrated in other animals.
An important consequence of this caching behavior is its vulnerability to exploitation by others. Many corvids observe conspecifics caching and are able to remember the precise location of the cache even days later. This has naturally led to counter-strategies in many species, such as caching behind barriers, waiting until no one is looking, pretending to cache, or decoy caching with inedible items (Heinrich & Pepper 1998; Bugnyar & Kotrschal 2002). Some birds return alone to a cache made in public in order to re-cache it in a more secure location (Emery & Clayton 2001). In this amazing experiment, hand-reared Western scrub-jays were allowed to cache either in private or while a conspecific was watching. Then they were allowed to recover their caches in private. It was found that individuals who had themselves stolen another birds caches in the past recached food in new sites only when they had been observed caching in the first place. Birds who had no experience in stealing caches did not attempt to relocate their caches. It seems that jays relate information about their previous experience as a thief of caches to the possibility of future theft by another individual and modify their recovery strategy by relocating caches. That is, these birds seem to, in effect, simulate another's viewpoint (an ability known as "theory of mind"). In a similar experiment Dally et al. (2004) gave scrub-jays the opportunity to cache in two trays, one that was located in an well-lit areas of the enclosure and the other in a darkened part of the cage. When observed by a conspecific, more caches were made in the darkened tray. However there was no preference for either the bright or dark tray when caching in private. The jays selectively recached items from the well-lit tray during recovery. In ravens, an experiment was conducted where a subordinate raven that had visual access to the location of hidden food led a dominant raven away from the food, before attempting to access the food themselves (Bugnyar & Kotrschal 2004). These examples are all highly suggestive of social reasoning in the form of tactical deception and theory of mind.
Concept of Number: The "Counting Crows"
Some of the earliest and most experimentally rigid tests of numerical concept in birds were done by Koehler in the 1950s with ravens (Corvus corax), jackdaws(Corvus monedulaI) and African grey parrots (Psittacus erithacus). He made use of two methods: simultaneous and successive presentation tests. Using the first, he presented the raven and the parrot with a card that had a number of dots drawn on it, and two small boxes-- one containing the correct number of items, and the other containing either one more or one less item than the correct number. He controlled for the birds discriminating based on quantity (and not number) by using two equally sized balls of modelling clay, one for each box, which were then molded into their respective number of items. Both birds rapidly learned to open the box with the correct number of items. In successive presentation tests, birds were rewarded for taking lids off of opaque boxes containing food items until a specific number of food items had been retrieved important feature of this experiment was the successive trials. That is, if the bird was rewarded for eating 3 items and there are 5 pots to choose from. For example, the bird would have to open only one pot if the first pot contained 3 items, but all 5 pots if only one item was in the first pot, 2 were in the fifth pot, and all the others were empty. jackdaws in this study stopped opening boxes after the correct number was eaten. Even more impressively, a jackdaw was able to master multiple forms of this task at once: the bird could open black lids until it had eaten two pieces of food, and stop; green lids until three food pieces, and stop; red lids until four pieces and white lids until five pieces (Koehler 1950). More recent work with numerical concept in crows corroborates these older findings (Smirnova et al. 2000), and similar studies have been done with chimpanzees and have seen comparable results (Boysen & Berntson 1989).
In Sum
In the foregoing mechanistic discussion of complex cognition in Corvidae, birds make adroit use of every tool in Emery's cognitive toolkit, the fact that our consideration is limited to the existing literature notwithstanding. In the above, causal reasoning and flexibility (innovation and insight) have been demonstrated by New Caledonian crows, rooks, and ravens; prospective and retrospective imagination by ravens and western scrub-jays. In fact, several species in the genus Corvus, such as C. corax, are known to use all of them. Corvids have been shown to have some concept of number and to exhibit self-awareness in mirror tests. These are sure signs of intelligence by almost any definition.
This website was created by Nathaniel Raley for Suzy Renn's Animal Behavior course (BIO 342) at Reed College, Fall 2010