Parasitic Behaviors of the Lancet Liver Fluke
Reed College - Biology 342 - Fall 2010
Genevra M. Kuziel
Phylogeny
Phylogenetic History of the Lancet Liver Fluke
The lancet liver fluke belongs to the phylum Platyhelminthes. Traditionally, this phylum is divided into three entirely parasitic classes, Cestoda, Monogenea, and Trematoda, and one mostly non-parasitic predatory class, Turbellaria. This organism is part of the class Trematoda (commonly referred to as the flukes) in which there are estimated to be between 18,000 and 24,000 species [8], almost all of which are obligate parasites (cannot live independent of a host) of molluscs and vertebrates.
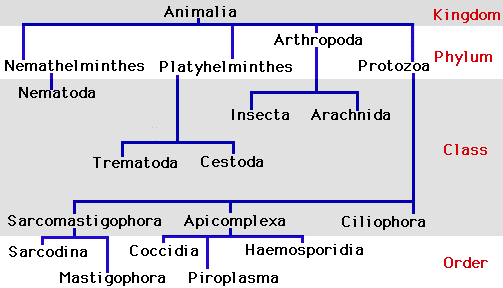
- Order: Plagiorchiida
- Family: Dicrocoeliidae
- Genus: Dicrocoelium
- Species: D. dendriticum
Two Possible Explanations for the Infection of Molluscs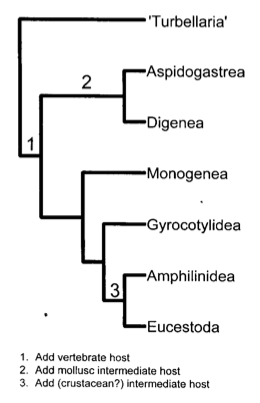
The class Trematoda is divided into two subclasses: Aspidogastrea and Digenea. Aspidogastreans are thought to be archaic trematodes and have a simpler life cycle, which most of the time includes mollusc and vertebrate hosts. Digeneans (which include the lancet liver flukes) are much more advanced trematodes. While the evolutionary origins of Digenea are still under debate, one hypothesis is that the proto-digenean was a parasite of a mollusc. Evidence for this comes from the fact that all digeneans have a mollusc as their first intermediate host, and as mentioned before, most aspidogastreans also parasitize molluscs. It might have been that the life cycles evolved through a gradual process of terminal addition when digeneans continued to survive predation of their host molluscs, eventually leading to the addition of other hosts until the complexity of the most recent life cycles developed. [4]
Figure 1. [1] A very simple phylogenetic history of the adoption of intermediate and definitive hosts. This diagram demonstrates an alternate hypothesis in which trematodes were parasites of vertebrates before they were parasites of molluscs. In this history Digenea and Aspidogastrea do still share the common trait of infecting molluscs at some point in their life cycles.
Evolutionary Foundations of the Host Manipulation Behavior
It is hypothesized that the sophisticated ability of the lancet liver fluke to use the mechanism of host manipulation to change the phenotype of its second intermediate host (see Mechanism for a more in depth explanation of this process) is a product of natural selection inherited from a common ancestor. The mechanism of host manipulation greatly increases the likelihood that the lancet liver fluke will infect its third definitive host. Examining this from an evolutionary standpoint, host manipulation is a wonderful example of the biological concept of extended phenotype proposed by Richard Dawkins. This concept states that the genes of one organism, such as those of the lancet liver fluke, can have phenotypic effects on another organism, such as the ant [10].
However, there are arguments that maintain that some changes in host behavior are actually not adaptive for the parasite but instead are merely by-products of infection. One such argument states that changes in the phenotype of a host result as "fortuitous payoffs of other adaptations" [10]. In the lancet liver fluke, a parasite that manipulates the behavior of its second intermediate host by encysting itself in the central nervous system, there is a possibility that the reason this location was initially favored was because it provided more protection from the immune system of the host ant than most other places in the host's body.
Still, this phenotypic change is so complex and so specific to the transmission of the lancet liver fluke that it seems like it couldn't be anything other than an adaptive manipulation, but until more is known about the complexity of the mechanisms used by the lancet liver fluke to alter the behavior of the ant, there is no real way to be positive. This is problematic because the fact is that it is still very hard to experimentally distinguish between an advantageous by-product and an advantageous direct product of natural selection [7]. Even so, it is likely that this parasitic trait evolved from a different function that happened to have beneficial side effects and improved the ability of the lancet liver fluke to be transmitted. As a result this host manipulation behavior improved the overall fitness of the lancet liver fluke, causing natural selection to favor and maintain this strategy in which the benefits of manipulation greatly outweigh the costs [7]. It may be that through this process the mechanisms gained a specific purpose and the behavior of host manipulation became a true adaptation.
In Platyhelminthes it appears that host manipulation has evolved independently more than once. Host manipulation has been documented in multiple families in this phylum, but they are often phylogenetically distant. One conservative estimate suggests that there are at least 20 separate cases in which the ability to manipulate host behavior has evolved among parasite lineages since life began on Earth [7]. Parasites face similar obstacles to transmission, a fact that has led to the evolution of similar adaptations to these problems (such as host manipulation) and similar life cycles in a broad range of parasite lineages. The wide variety of transmission routes where the successful transmission of parasites is determined, at least in part, by the behavior of the host suggests that the adaptive strategy of host manipulation has evolved repeatedly in many different lineages of parasites [7].
World Distribution of the Lancet Liver Fluke
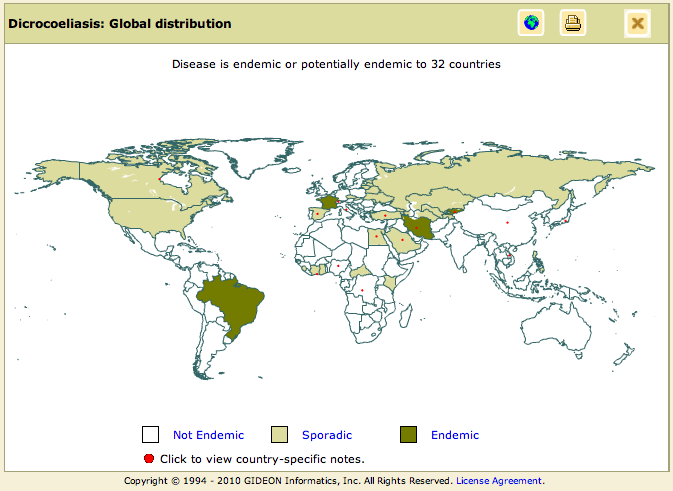
Note: Dicrocoeliasis is caused by both Dicrocoelium dendriticum and Dicrocoelium hospes.