THE IMMORTAL JELLYFISH
GEOFF DERVEN / BIOLOGY 342 / FALL 2010
| HOME | PHYLOGENY | ONTOGENY | MECHANISM | ADAPTIVE | REFERENCES | MORE? |
ADAPTIVE VALUE
Adaptive value considers how the function of a particular trait or behavior provides its organism with evolutionary benefit. Following Darwin’s example, all forms of life are challenged for survival by their very being-in-the-world. Whatever aspects prove to serve the greatest chance for success will be “naturally selected” for – these organisms are most likely able to continue on and reproduce (just as those who fail will perish). Adaptive value is thus perhaps the most straightforward means through which one can approach the study of a particular behavior: that is, put most simply, how is something beneficial to the animal?
At the same time, such a question becomes even more direct in its answer when dealing with T. nutricula, as the behavior at hand is one of cheating death. What could be more beneficial to one’s survival than not dying?
The greater life cycle of hydrozoan is typically reduced to two central stages – the polyp and the medusae. Bavestrello, et al. (1992) identified this sort of progression as already an adaptive strategy in itself; organisms capable of passing through multiple morphologies are more flexible at fitting into the changing particulars of their environment. Rather, both of these stages operate in entirely different modes of being, requiring their own distinct needs and benefits for proper survival and reproduction (Schmid, 1974). As T. nutricula do not subscribe to a unidirectional life cycle, they receive even greater adaptive benefit through bypassing this dichotomy, alternating between the most larval and mature modalities of life as necessary.
Piraino, et al. (1996) gathered a series of T. nutricula medusae and subjected them to various traumas throughout maturation. By starvation, radical changes in water temperature and salinity, and even physical mutilations via forceps and scissors, all of the T. nutricula that Piraino, et al. (1996) tested were transformed into stolons and polyps, reversing their lifecycle to revert to an earlier stage of existence – thereby withstanding all injuries. They would then resume their original, forward motion of development a second, third, or fourth time over. The ability to re-orient their life cycle in both directions creates a two-tiered rotation, allowing T. nutricula to adapt and recover from virtually any external threat.
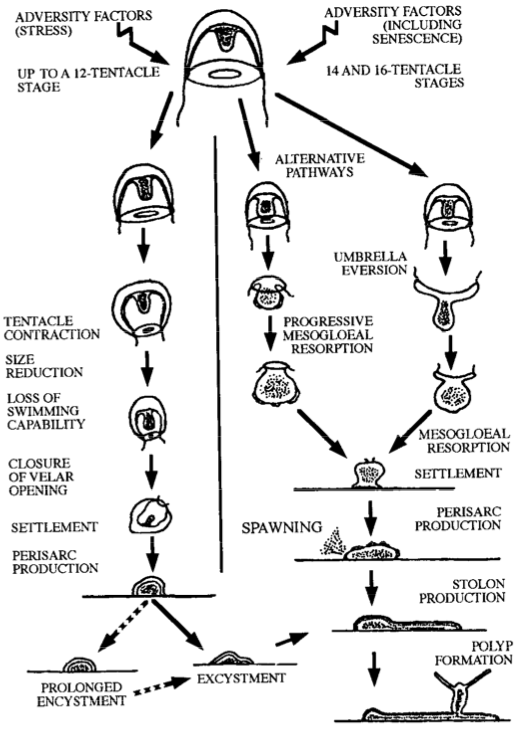
Potential pathways for transdifferentiation in Turritopsis nutricula (Piraino, et al., 1996).
Piraino, et al. (1996) claim that this immortalizing continuity could lead to an unending self-proliferation of T. nutricula. Miglietta & Lessios (2009) warn of the ecological disasters this could cause, especially when coupled with the usual hazards of invasive species – T. nutricula will inevitably overrun the habitats of less successful organisms. In fact, they are already reportedly spreading quite rapidly, via the ballast water of transoceanic ships (Hongbao Ma, 2010).