 |
High FidelityMonogamous behavior in mammalsMarjorie Nicholson & Kelsey Wood |
 |
Phylogeny
How did monogamy evolve?
Monogamy has evolved many times across the animal kingdom. There are monogamous species of insects, crustaceans, fish, birds, and mammals. While there is not a universal mechanism for monogamy across animals, it appears to evolve when the right environmental conditions make it adaptive.
Genetic differences within the rodent family (which includes Microtus), may provide clues to the phylogenetic history of monogamy.
Familia Rodentia: Primary Models In Mammalian Monogamy
Monogamous rodents include: House Mice (Mus musculus), Beavers (Castor family), Djungarian hamsters (Phodopus campbelli), Siberian dwarf hamsters (Phodopus sungorus), Prairie voles (Microtus ochrgaster), Pine voles (Microtus pinetorum).
 |
 |
The house mouse is monogamous. Image: DK Images. |
The nutria is polygynous. Image: C.C. Lockwood. |
{kind=link}
Some polygamous rodents: Rats (Rattus), Nutria (Myocastor coypus), porcupines, squirrels, gophers, montane voles (Microtus montanus), meadow voles (Microtus pennsylvanicus).
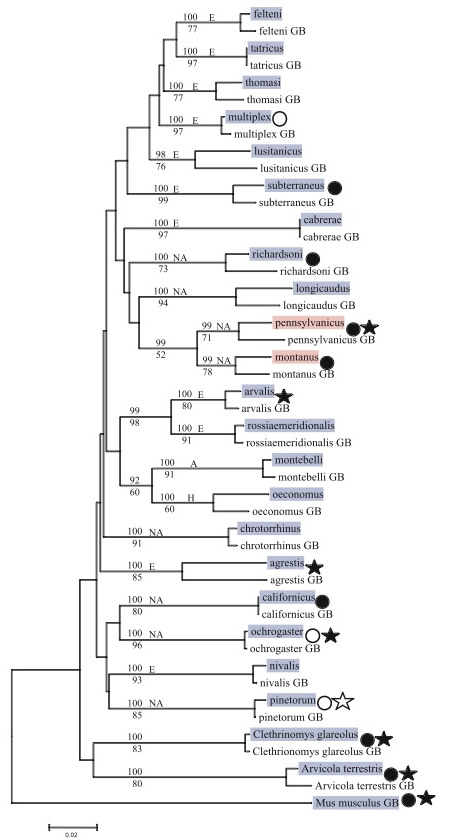 |
| Figure 1 from Fink et al. Black represents non-monogamy, white represents monogamy. Stars represent genetic (sexual) monogamy, white circles represent social monogamy. Blue highlights species with the long microsatellite, while pink highlights the short microsatellite. As you can see, many nonmonogamous rodents have the long microsatellite. |
V1aR expression in monogamous vs polygamous voles
Lim et al. (2004) notes that there are differences in the distribution of the vasopressin 1a receptor (V1aR) in the brains of monogamous and polygamous voles (see Mechanism for more on the function of this receptor) such that no two species have identical V1aR distributions. Lim et al. hypothesize that this phylogenetic plasticity in V1aR distributions could result in plastic behavioral responses to vasopressin, allowing monogamy to evolve when the environmental conditions are right.
Vole Genetics
Genetic analysis of the avpr1a gene (the V1aR coding region) shows 99% conservation of gene sequence between vole species, but monogamous prairie and pine voles have a longer microsatellite region that the promiscuous montane and meadow voles (Lim et al, 2004). The longer microsatellite for avpr1a is associated with an elevated level of gene expression. However, Fink et al. (2006) looked at a larger sample of rodent species and could not find a link between microsatellite length and mating system (see figure). Their analysis indicates that the long microsatellite region is the ancestral state. They also point out that mammalian monogamy could not be determined solely by the single gene avpr1a because we know that pair-bond formation is dependent not only on vasopressin, but also on oxytocin and dopamine (see Mechanism). Thus the differences between species are probably due to changes in multiple genes that code for vasopressin, oxytocin, and dopamine receptors.
West-Eberhardian Evolution
In West-Eberhard’s theory of evolution, a new trait arises because of developmental plasticity, and then genetic accommodation may or may not follow. Within the rodent family, it is possible that monogamy appeared in prairie voles due to changing environment, and the longer microsatellite region simply helped facilitate and reinforce monogamy after it appeared.