
|
Bipedal Locomotion in LizardsBiology 342 Fall 06 |
Mechanism
How do these lizards actually accomplish bipedal locomotion?
In 1949, Richard Snyder set out to determine how lizards
execute bipedal motion. To this end, he took high speed videos (64
frames per second) of Basiliscus basiliscus exhibiting the behavior and
analyzed the movement from three different perspectives. Snyder was
able to conclude that the forelimbs do not play an important role in
elevating the anterior section of the torso, but that the epaxial
muscle in the back allows for this maneuver. Snyder observed that
Basiliscus can begin bipedal motion from a resting position with a
thrust by both hindlimbs of the animal followed by one hindlimb rapidly
swinging around in a lateral arc to catch and support the weight of the
body and take the first bipedal step. It has also been observed that
Basiliscus can commence bipedal motion from quadrupedal motion by
raising the anterior portion of the torso and head. Snyder presents the
diagram shown below of the physical stages of bipedal motion.  Snyder notes that the proximal portion of the elongated tail of
Basiliscus is elevated during bipedal motion, although the tip of the
tail may drag. He also determines that the tail acts as a
counterbalance to the trunk and head of the body. Snyder tested his
theory by removing portions of the tail of a Basiliscus, allowing it to
acclimate, and inducing bipedalism. As the length of the tail is
shortened, the ability of the lizard to execute bipedal locomotion is
negatively effected. Sufficiently shortening the tail can prevent a
lizard from being able to move bipedally.
Snyder notes that the proximal portion of the elongated tail of
Basiliscus is elevated during bipedal motion, although the tip of the
tail may drag. He also determines that the tail acts as a
counterbalance to the trunk and head of the body. Snyder tested his
theory by removing portions of the tail of a Basiliscus, allowing it to
acclimate, and inducing bipedalism. As the length of the tail is
shortened, the ability of the lizard to execute bipedal locomotion is
negatively effected. Sufficiently shortening the tail can prevent a
lizard from being able to move bipedally.
Joshua Laerm performed another experiment using Basiliscus basiliscus in 1973. In this experiment, Laerm set out to determine any differences in bipedal mechanics on different substrates. High-speed motion pictures were taken (300 frames per second) of lizard bipedalism in water and on a sandpaper-covered platform. From these recordings, he was able to determine that there is no difference in the basic motion of bipedalism based on substrate. There are, however, observable differences in terrestrial and aquatic bipedalism. He explains that the lateral undulation of the spine is exaggerated in the aquatic form of this motion. The reason for this, they explain, is to counter drag in the water. There is also a difference in angle of trunk incline from a range of 25-35 in water to 20-46 degrees on land.
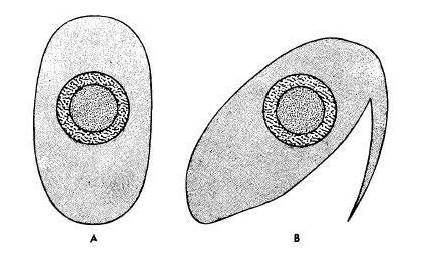 Laerm also described a special scale on the Basilisk's toe that helps
it run across water. The image at left, taken from Laerm's 1973
publication, shows a cross section drawing of the second phalange of
the third toe in an iguana (A) and a basilisk (B). This special scale
is visible here as a fringe protruding from the ventral lateral side of
the toe. The image below shows how this structure functions on
terrestrial substrate (B) and on water (C).
Laerm also described a special scale on the Basilisk's toe that helps
it run across water. The image at left, taken from Laerm's 1973
publication, shows a cross section drawing of the second phalange of
the third toe in an iguana (A) and a basilisk (B). This special scale
is visible here as a fringe protruding from the ventral lateral side of
the toe. The image below shows how this structure functions on
terrestrial substrate (B) and on water (C). 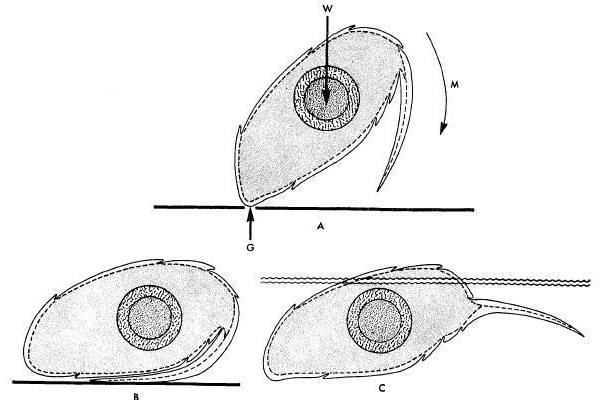 By flaring or compressing this fringe, the lizard is able to travel
adeptly over land and water. When the fringe is removed, more of the
lizard's body is below the water line, but aquatic bipedal motion is
still possible. From this, Laerm is able to conclude that the fringe
aids in vertical support of the lizard in the water and in locomotive
efficiency, but is not necessary.
By flaring or compressing this fringe, the lizard is able to travel
adeptly over land and water. When the fringe is removed, more of the
lizard's body is below the water line, but aquatic bipedal motion is
still possible. From this, Laerm is able to conclude that the fringe
aids in vertical support of the lizard in the water and in locomotive
efficiency, but is not necessary.