Exam 1
1.Reinforcement will result in the evolution of pre-zygotic isolating barriers, and will arise when hybrids are being formed and when those hybrids have lower fitness. This condition favors the evolution of pre-zygotic traits that prevent hybrids from forming in the first place.
for each trait that follows, the possible role for reinforcement is indicated in parentheses (YES/NO).
The identified reproductive isolating barriers for milkweeds include: flowering time (YES), ecological/habitat (POSSIBLY), mechanical- pollinia size and stigmatic chamber size (YES), and some form of post-mating barrier that could be either a pre-zygotic incompatibility mechanism (YES) or a post-zygotic effect like hybrid inviability (NO). An ethological barrier involving pollinator foraging behavior was evident, but it appeared that pollinator species were mostly shared among the milkweeds. However, a given pollinator often restrcited its visits to one species on each foraging bout -- if the floral constancy is due to plant traits that have diverged (Allowing species distinctions to be made), the plant traits could have evolved through the Wallace Effect (YES).
2. Populations in allopatry and without connection through gene flow are expected to diverge genetically over time. This will happen because of the inevitable effects of mutation, coupled with genetic drift and natural selection. For the following description, this divergence need not involve traits that are , themselves, invovleed in pre-zygotic reproductive isolation. Epistasis describes interactions between loci, such that effects of the genotype from one locus depend on the other loci they are with (i.e the phenotypic effect of a genotype at one locus depends on the genetic background). Often epistatic interactions can reduce fitness. Alleles that produce negative epistatic effects will not arise and spread to fixation in a genetic background in which they are disfavored by selection. If epistasis is a general feature of genomes and governs the way genes interact with each other, we can expect that any genetic divergence over time among allopatric populations will lead to the accumulation of Dobzhansky-Muller-Bateson incompatibilities. These interacting loci will contribute to post-zygotic reproductive isolation only when the two formerly allopatric populations meet and interbreed; at that time, epistatic effects of the DMB loci are expressed in the hybrid offspring. The consequence of genetic divergence (whether caused by local adaptation or genetic drift) will therefore inevitably lead to the inability of populations to interbreed successfully as a result of post-zygotic isolation. Ifnot in allopatry and gene flow is occurring between the two populations, their genetic backgrounds are less likely to diverge, and this lessens the opportunity for DMB loci to diverge (since the alleles with the epistatic effects on fitness are being "tested", by selection, in the same genetic background in both populations)
3. For Stephanomeria, the allopatric model could involve a range expansion for S. exigua into Oregon at the northern margin of its range involving a very small number of colonists (founder event), or migration of a large population followed by a population collapse (bottleneck) in Oregon. As a result of the small population size for several generations, the Oregon population evolved differences (including chromosomal rearrangements and self-compatibility) that provided reproductive isolation form the California S. exigua. This is the "peripatric" variant of the allopatric mode of speciation, with an important role for genetic drift. When S. exigua migrated once again to Oregon, it encountered the diverged population from which it was now reproductively isolated. Gottlieb's data from the current geographic distribution seem to be consistent with either the allopatric or sympatric scenario. However, the allopatric model's local extinction of S. exigua and then migration back into the Oregon site may seem implausible. Also given the seed bank that Gottlieb describe for the species, it may be hard to envision that S. exigua was ever totally gone form the Oregon site once it first arrived.
For Howea, the allopatric scenario would require at least two migration events to the island (which is arguably too remote to make this plausible). Under the allopatric model, either the two species evolved elsewhere as separate species in allopatry and arrived to the island in independent migration events, or one species arrived on the island, evolved into the second species (and diverged for soil and flowering time traits), and then a second migration of the first species to the island resulted in their current sympatry. The first scenario seems implausible because of the extreme remoteness of the island and because the two species are not found anywhere else but Lord Howe Island. They are also each others closest relatives (sister taxa) based on phylogenetic analysis. The second scenario also seems implausible given the remoteness of the islands. The timing of the speciation event could have refuted the sympatric hypothesis if the phylogenetic estimate (based on a molecular clock) showed the species to be older than the geologic age of the island, but instead, the speciation node on the phylogeny for the two sister taxa was estimated to be younger than the island's age.
4. various answers/justification possible.
YES – Mayr examined the kinds of taxa for which the BSC has genuine difficulties (asexual reproduction, hybridization, taxa with various ploidies that make interbreeding difficult to judge) and found that these taxa reflected the minority of species in the Concord Flora, thereby showing that botanists' concerns with the BSC are not warranted.
NO – Mayr applied a definition that invokes reproductive isolation and interbreeding without any experimental support from performing empirical tests of interbreeding. He used morphology in many cases to infer the degree of interbreeding. He tackled the species question in a setting without concern for spatial variation (the extent to which interbreeding occurs for geographically distinct populations of a single species) and without concern for temporal variation. His study also focused on a flora that is extremely well-studied, although it’s not clear if this would lead to a biased conclusion about species definitions (i.e., does study of a flora minimize the occurrence of asexual reproduction and hybridization or other problems for the BSC?). It might also be argued that, given his methods, a well-studied flora was a requirement for the study he conducted.
5. Mechanical and Ethological are the most likely current RI barriers (the question suggests they RI barriers must be pre-mating barriers, and temporal and geographic are ruled out since they are in the same communities and flowering at the same time).
These barriers most likely arose in allopatric populations of a single species, where selection by the local pollinator species (differing between populations) resulted in local adaptation to that pollinator and consequently ethological and mechanical features of the flower that aid in pollination by the pollinator that is specific to that local habitat. When these orchid populations have subsequently come back into sympatry (secondary contact), they are now isolated species by virtue of the mechanical/ethological changes in their floral traits that resulted from local adaptation in allopatry.
(NOTE: the evolution of mechanical/ethological barriers in sympatry is much less likely to occur because of the difficulty of splitting a single, randomly mating population into two isolated populations through disruptive selection that focuses on adaptation to different pollinators. If a new mutation that makes an individual utilize a new pollinator confers an advantage, the entire population will shift to that new pollinator. If the new mutation is disadvantageous, it will be lost. You are unlikely to end up with two isolated populations in sympatry using different pollinators if the entire process happens in sympatry).
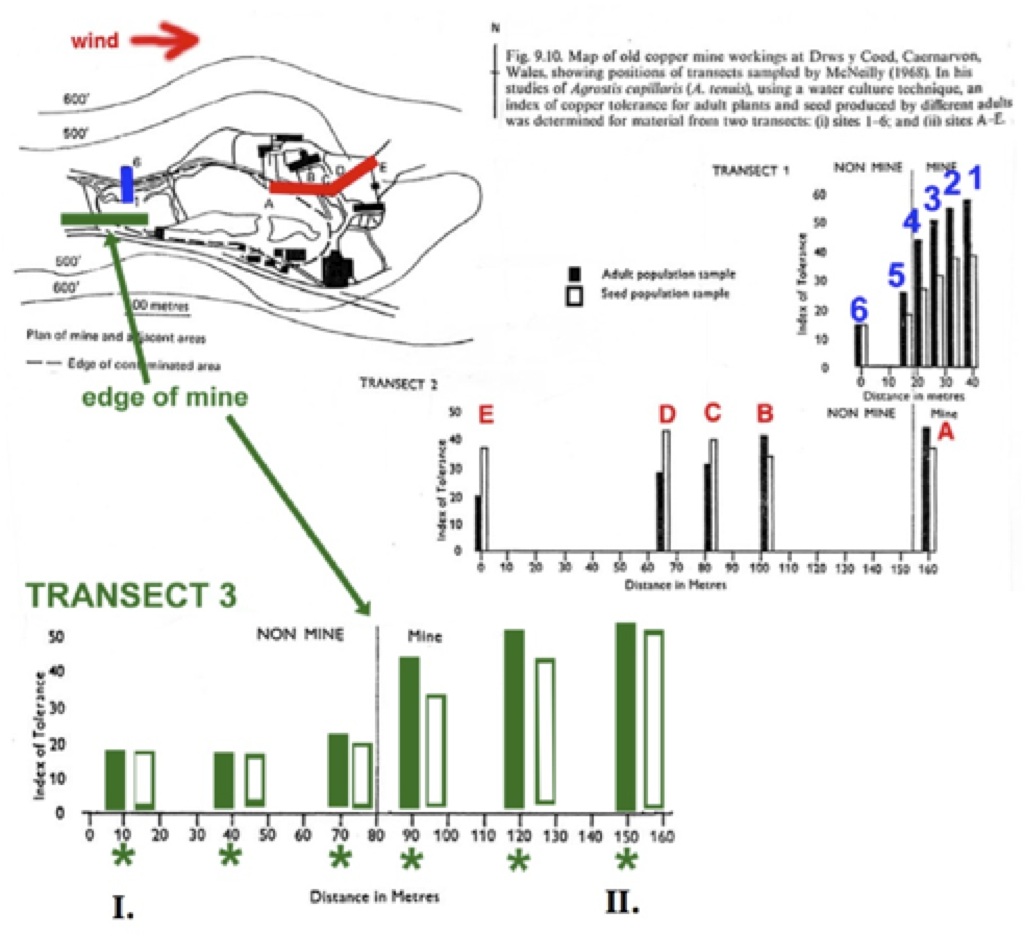 6. Natural selection off the mine (position I) favors low copper tolerance (because of the competitive advantage for non-tolerant plants). On the mine (position II), selection favors copper tolerance in the plants. The other factor important in this transect is the influence of gene flow, which is predominantly occurring from off the mine towards the mine. The offspring of parents at any of the six sampling sites will reflect the effects of gene flow, which in most cases are carrying "non-tolerance" alleles, so the offspring will be generally as tolerant (position I) or less tolerant (position II) when compared to the parents.
6. Natural selection off the mine (position I) favors low copper tolerance (because of the competitive advantage for non-tolerant plants). On the mine (position II), selection favors copper tolerance in the plants. The other factor important in this transect is the influence of gene flow, which is predominantly occurring from off the mine towards the mine. The offspring of parents at any of the six sampling sites will reflect the effects of gene flow, which in most cases are carrying "non-tolerance" alleles, so the offspring will be generally as tolerant (position I) or less tolerant (position II) when compared to the parents.
The first two sampling sites (off the mine) show adults that are non-tolerant (therefore locally adapted) and offspring that reflect gene flow from other non-tolerant individuals. The third site near the mine border might show increased CU tolerance in the adults as a result of selection to match increased levels of copper that may be in the soil near the mine's edge, but their offspring will have less tolerance due to gene flow from the non-tolerant individuals upwind and off the mine. For the three sampling sites on the mine, the tolerance shown in the adults will be high, while the offspring will be less tolerant due to gene flow from off the mine. The difference between parents and offspring will be greatest near the mine boundary where gene flow from off the mine is most effective. Further into the mine, there will be less gene flow from the non-tolerant plants off the mine which will result in both parents and offspring showing greater and more similar adaptation through copper tolerance to the mine's soils.
7. i.) SUPPORT: reinforcement should result in greater reproductive isolation between sympatric populations of two species and lesser isolation between more allopatric populations (which is the pattern observed). Note that reinforcement must entail pre-zygotic barriers to have resulted from selection for reinforcement, and these would be expected to reduce the actual formation of hybrids under natural conditions. The reduced seedset would therefore need to result from pre-zygotic stages of reproductive isolation (anything pre-mating, or post-mating but still prezygotic). If the failed seedset for the sympatric populations is due to hybrid inviability, this trait did not evolve through reinforcement
7 ii) this could be interpreted either direction:
REJECT: This appears to be the best interpretation, as the results demonstrate an opposite pattern from that predicted by the reinforcement hypothesis. For pollen germination, the sympatric populations have less pre-zygotic reproductive isolation than do the allopatric populations. This result would only show that reinforcement has not acted upon pollen germination (pre-zygotic, post-mating); reinforcement could still have affected on the evolution of other (pre-zygotic RI) traits.
SUPPORT: the results demonstrate that post-mating barriers are not adequate to isolate the two species, which might provide the necessary selective conditions for further reinforcement of premating isolating barriers to take place.
7 iii) The competition in the past for the resource (pollinators) must have involved the pink morphs of the two species, so the experiment would involve looking for evidence of pollinator discrimination when the pink morphs of the species are placed together (interspecific competition) compared to when they are alone (intraspecific competition). It might be best to do this utilizing the pollinator species available in the curent zone of sympatry (where the competition would likely have taken place). Given the asymmetry of the divergence, we might predict that P. drummondii will be the loser in the competitive interaction (since that is the species who's flower color diverged in the area of sympatry where past interspecies competition might have taken place).