Cross-Species Tool Use
Biology 342 Fall 2015
Jasmine Huang & Lauren Vanderhooft
Ontogeny
Observational and social learning
Observational or social learning is one way that behavior can develop over an organisms lifetime. A comprehensive definition of social learning states that the behavior (1) must be a learned behavior, (2) it must be acquired through some form of social transmission, often by watching or interacting with another individual, and (3) it must persist even in the absence of the demonstrator (Wynne & Udell, 2013 pp. 201-202).
Social learning can be categorized into two categories, horizontal and vertical transmission. Horizontal transmission refers to social learning between animals in the same peer group or generation, such as juveniles learning from juveniles. Vertical transmission describes transmission across generations, including transmission in both directions. For example, upwards transmission could be from offspring to parent, and downwards transmission could be from parent to offspring.
Studies introducing novel species of nuts to chimpanzees demonstrate that social learning influences the acquisition of tool use behavior, such that chimps will selectively choose to learn from older conspecifics (downwards transmission) or from same-age peers (horizontal transmission), but not from younger animals (upwards transmission) (Biro, Inoue-Nakamura, Tonooka, Yamakoshi, Sousa, & Matsuzawa, 2003). Social transmission of handedness in tool use has also been observed in chimps (Biro et al., 2003).
Another primate, orangutans, also demonstrate social transfer of tool use (Fox, Sitompul, & Van Schaik, 1999). When given a novel task requiring tool use in the field, orangutans in social groups transmitted tool using behavior more quickly than orangutans in less social groups, even though orangutans are naturally predisposed to sparse social groupings (Fox et al., 1999).
Juvenile Asian elephants learn to use tree branches to deter flies only after observing their mothers exhibit the behavior first, another example of downwards transmission (Hart, Hart, McCoy, & Sarath, 2001).

This juvenile elephant has learned to modify a tree branch from observational learning from older elephants in its peer group, as well as how to use the branch as a tool. (Image from Hart, Hart, McCoy, & Sarath (2001), p. 846, Fig. 3.)
In dolphins, the use of sea sponges as foraging tools is socially transmitted through imitation within and between generations of highly related animals, generally through the mother (Krützen, Mann, Heithaus, Connor, Bejder, & Sherwin, 2005).
This video simulation of social behavior between dolphins demonstrates the possible capacity of dolphins transmit tool-using behavior. (Video from NMANewsDirect on YouTube.)
Observational learning of tool use does not only occur in mammals. Another species that demonstrates social learning of tool-use behavior is the New Caledonian crow. These corvids can acquire tool manufacturing techniques from observing other animals, but do not require observation to develop the behavior, as they often spontaneously use tools (Bluff, Weir, Rutz, Wimpenny, & Kacelnik, 2007; Emery & Clayton, 2009).

This crow is using a stick to get food from a tree hollow. Behavioral testing demonstrates the crows can use an array of tools flexibly and break routine even when past experience has produced positive outcomes. (Image from DOGO News.)
The term cultural transmission has also been used to describe the tool use patterns in the New Caledonian crow. These animals natural habitat is the small island of New Caledonia, where field research has shown distinct pockets of animals that vary in the types of tools that they use, suggesting that there is a social component to the selection of tools (Bluff et al., 2007).
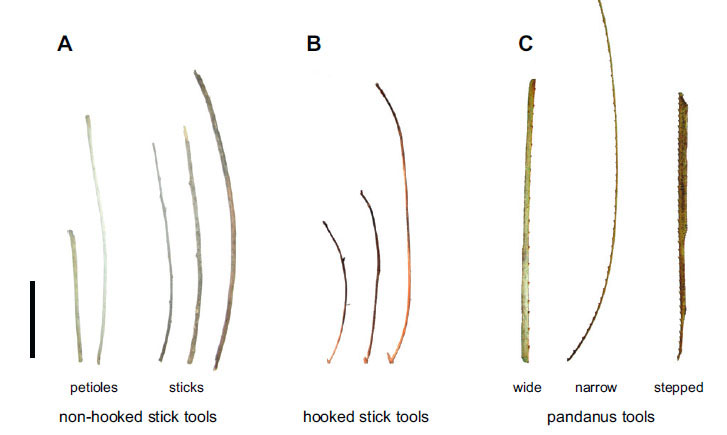
Crows have the capacity to use different styles of tools within the same tool class, as with these sticks that are both found (non-hook and hooked sticks) and modified (pandanus tools). (Image from Rutz & St Clair (2011), p. 155, Fig. 3.)
Critical period
In some species, there is a critical or sensitive period in an organisms life where tool use must be learned or the behavior will not develop, or not develop proficiently. In chimps that crack nuts, longitudinal data suggests that learning to use tools typically occurs between 3-5 years of age (Biro et al., 2003). For woodpecker finches, a species that does not spontaneously use tools in the wild, captive juveniles readily learn this behavior while captive adults are unable to acquire tool use (Tebbich, Taborsky, Fessl, & Blomqvist, 2001). These data suggest that there is a critical period where this behavior must be learned.
While this older chimp may not be intentionally performing a tool-using behavior to teach a young chimp, the young chimp may attend to the behavior nonetheless and learn through observation how to crack open nuts. (Video from purepedantry on YouTube.)
Trial-and-error learning
Some tool using species acquire tool use behavior from trial-and-error learning rather than social learning. In the critical period for tool use development in woodpecker finches, previous attempts at tool use inform the next attempt (Emery & Clayton, 2009; Tebbich et al., 2001).
Trial-and-error learning is inherently flexible to the circumstances. For example, rooks first use small rocks to displace water, raising water level to reach a worm at the surface before choosing larger rocks to displace more water in one movement; after experiencing the same set up with sawdust instead of water, rooks stop performing the act of placing pebbles in the sawdust (Bird & Emery, 2009).

This rook is using a heavy rock, as opposed to other smaller and lighter rocks, to open a trap door at the bottom of the tube to gain access to a worm. (Image from BBC News.)
Experience
An organisms previous experience with using tools can affect the degree of flexibility in tool use.
Capuchins with greater experience with sticks and tool tasks are less likely to perform flexible tool use behavior to achieve a desired outcome, possibly due to overgeneralization from a previous tool use task, indicating a lack of causal reasoning (Visalberghi & Limongelli, 1994). On the other hand, previous tool using attempts can successfully inform future attempts, as with woodpecker finches (Emery & Clayton, 2009; Tebbich et al., 2001).

This woodpecker finch, like crows, is able to use sticks to access food in tree hollows. (Image from Animals Ekstrax.)
Brown bears have been shown to engage in crude tool use by manipulating stones with barnacles to scratch themselves after foraging and during molting season (Deecke, 2012). This is comparable to how the bears regularly rub against stationary rocks, but the stone as a tool acts to immediately serve feedback that does not require developmental learning.

These time frames show a brown bear manipulating a stone in a scrubbing motion, possibly to clean its muzzle or scratch itself. (Image from Deecke (2012), p. 728, Fig. 1.)
Wild or Captive
In some observed cases, tool use develops differently in a single species due to the environmental differences between captivity and the wild. Orangutans in the wild use found objects as tools, while captive orangutans have the capacity for more flexible tool use (Fox et al., 1999).
Captive elephants engage in fly swatting behavior by using branches as tools. In the wild, elephants are not known to use branches for this purpose (Hart et al., 2001).
Two corvid species, rooks and jays, do not use tools in the wild, but when presented with food retrieval tasks in captivity requiring tool use, they can aquire this behavior (Bird & Emery, 2009, Cheke et al. 2011). This highlights the importance of environmental differences on the development of tool use behavior.