Eusocial Behavior
Biology 342 Fall 2014
Karl Menzel and Allie Buckner
The Evolution of Eusocial Behavior
How Do Species Become Eusocial?
 There are several theories proposed by entomologist Charles D. Michener on how eusocial behavior developed. Michener dubbed two routes to eusociality parasocial and subsocial.
There are several theories proposed by entomologist Charles D. Michener on how eusocial behavior developed. Michener dubbed two routes to eusociality parasocial and subsocial.
Parasocial behavior begins with the adults of one generation living together in one nest or area. Each adult takes care of its own young. This behavior can be seen in Sociable Weaver birds, who work together to create massive communal nests, but each takes care of their needs. (Picture of a Social Weaver nest in a telephone pole on left.) After species evolve to live communally, they develop quasi-social behavior.
Quasi-social behavior is characterised by the community working together to care for young.
Finally, species who are on their way to eusociality will become semisocial, which is characterized by a development of castes and a division of labor. This is the stage when a sterile caste will emerge from fertile animals. Animals will be considered eusocial only when multiple generations live together in their communal nest held together with a strong caste system.
Michener has proposed one other route to eusociality, called subsocial. Instead of many members of a species becoming eusocial together and developing a caste system from social interactions, subsocial animals are solitary. This means that a fertile female will live separately from other members of a species and she will give birth to offspring that she will take care of alone. This lifestyle will become eusocial when she lives long enough to have many generations of offspring and her offspring develop into different, sterile castes with different jobs
Hamilton’s Rule
Why would an animal ever be altruistic? If an evolution selects for animals with high lifetime reproductive fitness then how could an altruistic behavior ever evolve? The answer has to do with the success the animals have of passing on their genes. This makes sense for an individual, a gene that allows an individual to produce more successful offspring will get passed on in those offspring and be maintained in the species. This relationship also explains how it would evolve for an animal to be altruistic. Hamilton's rule dictates that an individual will risk detriment to themselves to help another as long as the amount of genetic material that the two individuals share outweighs the cost. This leads us to Hamilton’s rule: r B > c in which r is the relatedness of the two animals, c is the cost of the action, and B is any other benefit that the animal might receive from doing the action. So if a gene codes for a behavior that harms the animal that does it, but helps closely related animal, there is a possibility that the related animal also has the same gene. That is why animals seem to perform altruistic actions.
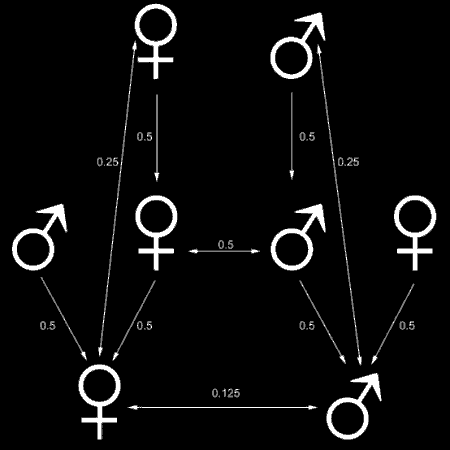 For haploid-diploid eusocial species like bees, worker’s children will be less related to them than their sisters. Female bees have two sets of chromosomes for passing genetic information (they are diploid), while males only have one (haploid.) Half of the daughter’s genes are her father’s and half her genes are her mother’s. The 50% of her father’s genes are shared by her sisters and 25% of her mother’s genes are shared by her sisters, that means that sisters have a relatedness of 75%.
For haploid-diploid eusocial species like bees, worker’s children will be less related to them than their sisters. Female bees have two sets of chromosomes for passing genetic information (they are diploid), while males only have one (haploid.) Half of the daughter’s genes are her father’s and half her genes are her mother’s. The 50% of her father’s genes are shared by her sisters and 25% of her mother’s genes are shared by her sisters, that means that sisters have a relatedness of 75%.
This is shown in the diagram to the left, where the top row is the queen and her male, the arrows show what portion of their genes they are passing and to which offspring. Horizontal arrows show relatedness.
If non-queen females were to have babies, they would only be 50% related, which is smaller than the relatedness between sisters. Hamilton’s Rule predicts behavior that benefits animals with higher relatedness. This kind of behavior is best seen in reproductive policing.
Sources used for this page in order of section:
.