Laughter in Chimpanzees and Humans - Biology 342 Fall 2012
By Yuan Xue, Soso
How is Laughter Evoked and Vocalized in Chimpanzees and Humans?
The stereotypic laughter behavior is induced by an ambivalent stimulus (such as tickling or a joke), evoking a mixture of pleasurable and unpleasurable feelings (Plessner, 1961), which leads to an involuntary stereotyped motor reaction. The neural processing of an external stimuli, whether it be auditory, tactile, visual, or a combination of these, in the brain leading to motor facial display and vocalization of sounds that we recognize as "laughter" is an an area of intense study.
Neural Circuitry of Laughter in Humans
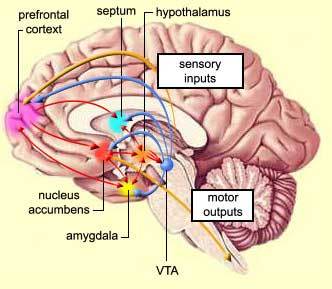
Based on classical understanding of neurology, most scientists accept that laughter elicitation begins at the transmittance of external signals through sensory nerves to somatosensory cortex or auditory cortex; the neural activity is then processed in prefrontal lobe for higher order decision making, and finally transmitted to motor cortex via the rest of limbic pathway for possible modulation of emotional responses. There is controversial debate on whether if laughter has other higher order regulatory system, and precisely what neural structures are responsible for such regulation.
The expression of laughter seems to depend on two partially independent neuronal pathways: the first being "involuntary" or "emotionally driven" system which involves amygdala, thalamic/hypo- and subthalamic areas and the dorsal/tegmental brainstem; the second being "voluntary" system which originates in the premotor/frontal opercular areas and leads through the motor cortex and pyramidal tract to the ventral brainstem (Wild et. al, 2003). The two systems and the laughter response appear to be coordinated by a laughter-coordinating center in the dorsal upper pons. Neurological studies on laughter pathways are conducted based on pathological reports of human patients with lesions and MRI/magnetic resonance angiography of neural activity during laughter.
The "voluntary" laughter system is dissociated from "involuntary" system in that the area of the mesencehalon and pons has a functional division between those structures necessary for the formation of "involuntary" laughter on the one hand, and of "voluntary" laughter on the other. Ventral lesions in these areas lead to pareses of volitionally created facial expression with either no damage to or exaggerated expression of "involuntary" driven laughter expressions. Wild et. al postulates that in a genuine, emotional laughter, the frontal and temporal regions of the brain are involved in the perception of sensory inputs which in turn would induce facial reactions and laughter mediated by dorsal brainstem regions. These reactions would be inhibited by frontal motor/premotor areas. Damages to this inhibitory system may result in pathological laughter (Psudobulbar affect, or PBA) in which pressure-induced disruption of inhibitory tracts results in forced facial expression. This notion that limbic system (thalamas, hypothalamas, hippocampus, amygdala, septuma and other associated strucutres) and motor cortical pathway together play a critical role in laughter is further supported by functional magnetic resonance imaging (FMRI) study conducted by Wattendorf et. al on human subjects and laughter.
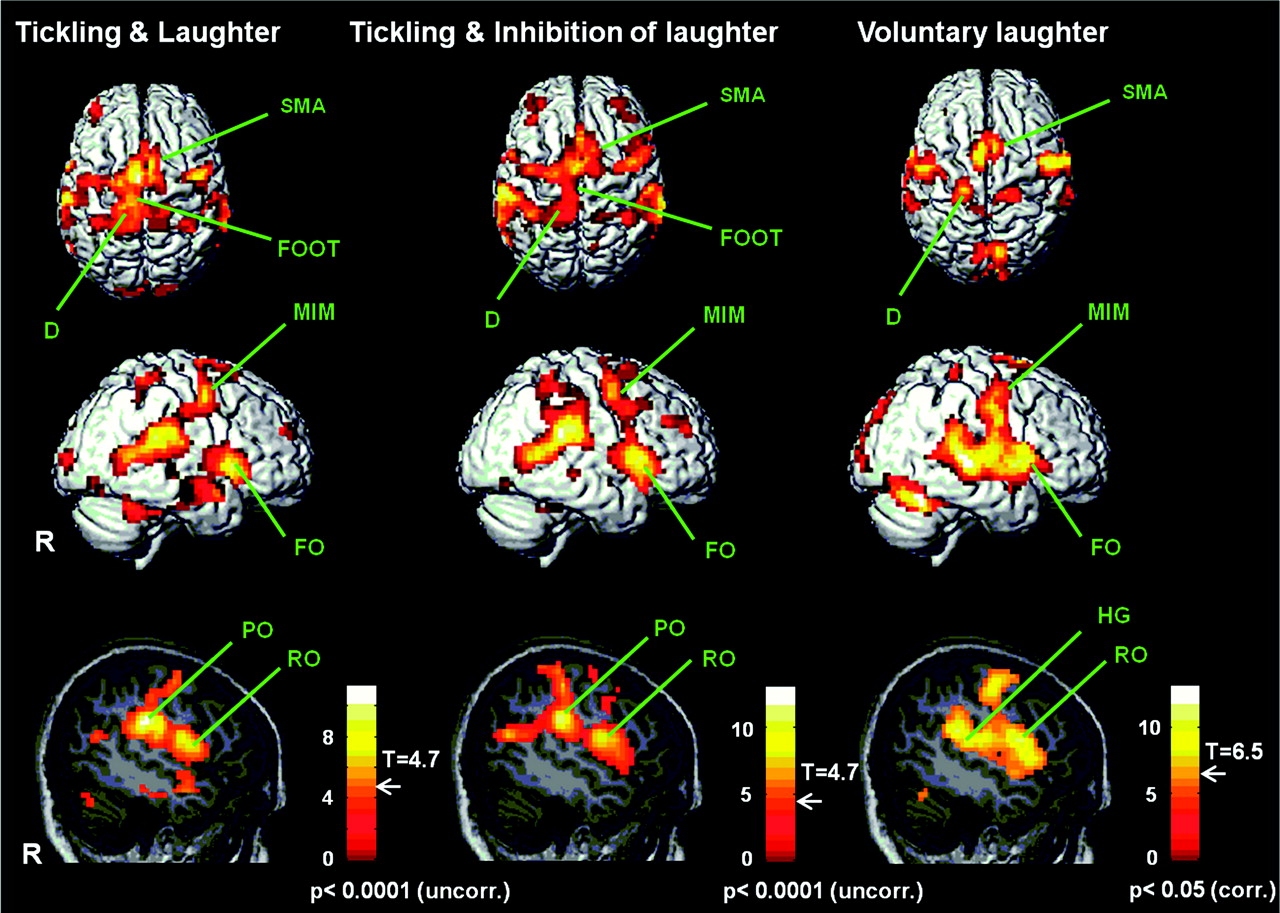
FMRI of human subjects in three treatment groups: Tickled on the right foot sole and allowed to laugh (T), Tickled on the right foot sole and asked to suppress laughter deliberately (I), and voluntary laughter without tickling (L). Three-dimensional top (first row) and lateral (from the right: R) views (second row) of the brain, and sagittal slices through the right hemisphere (third row). Activity is apparent in the primary sensorimotor areas (S1/M1) representing the diaphragm (D), mimic musculature and larynx (MIM), and the pharynx (Rolandic operculum: RO), as well as in the SMA and in the frontal operculum (FO). Activity in the left-foot area (FOOT) and in the parietal operculum (PO) is observed only during T and I. The auditory cortex (Heschl's gyrus: HG) is activated only during L. T: P < 0.0001 (uncorrected); I: P < 0.0001 (uncorrected); L: P < 0.05 (FWE-corrected). On each scale, the threshold level (T-value) for observed activity is indicated. (Wattendorf et. al, 2012).
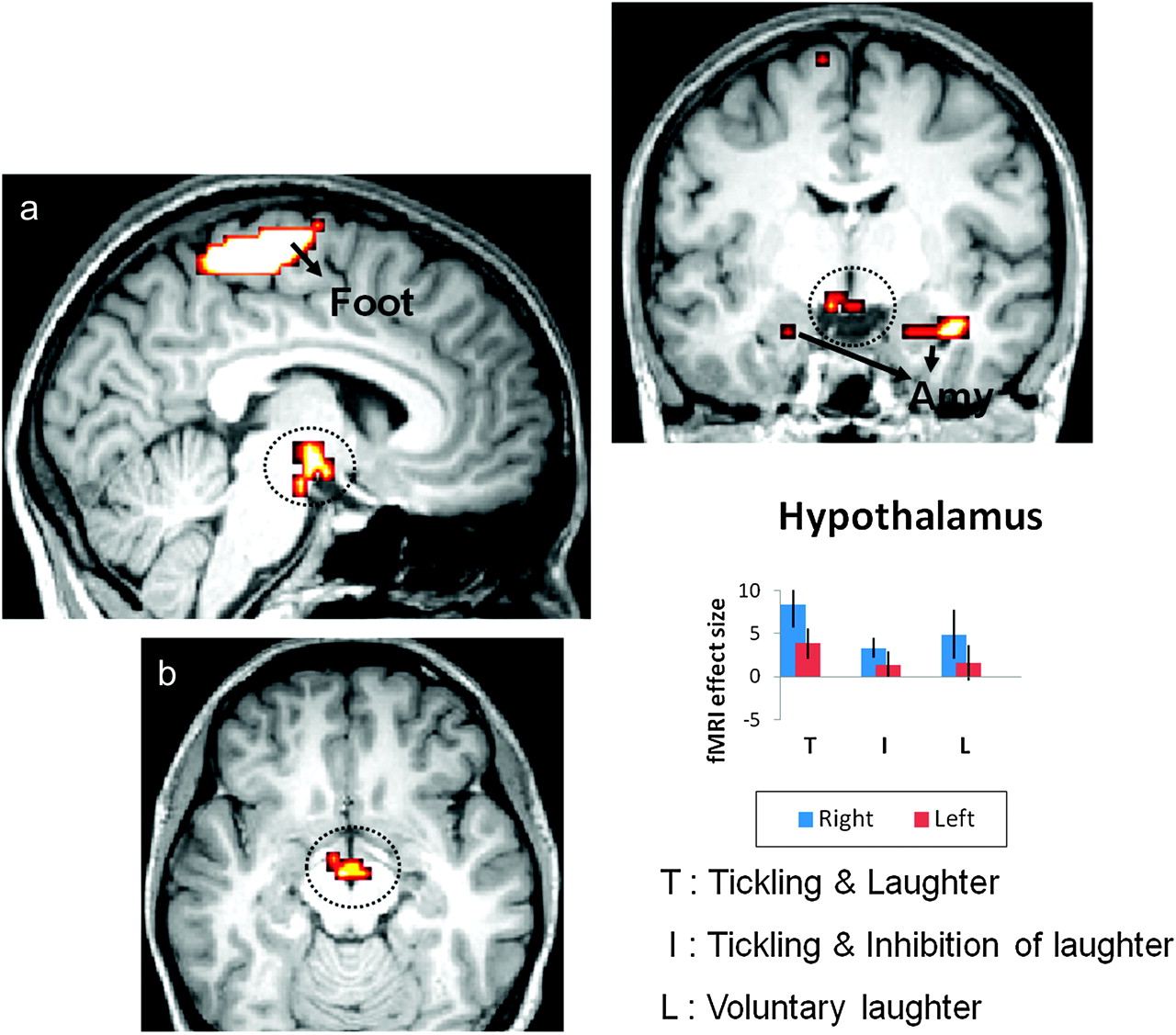
Conjunction analysis and effect size. Increased significance of hypothalamic activation in response to T versus I and versus L (a: sagittal paramedian; b: axial; c: frontal; P < 0.005 [uncorrected]). Parameter estimates (beta values) that were derived from the peaks of activity confirmed a higher level in this region during T than during either I or L (bar graph). Activity is also apparent in the amygdala (Amy) and in the somatosensory area representing the foot (Foot). The statistical maps are superimposed on an MNI-normalized image of the brain. Error bars indicate the 90% confidence interval. (Wattendorf et. al, 2012).
Wattendorf's FMRI studies shows that there is only hypothalamic activation in (T), whereas (I) and (L) did not activate hypothalamas, which confirms the hypothesis that the limbic pathway of vocalization is critically involved in laughter induced by tickling. Laughter is also associated with specific activities in higher order sensory-motor areas such as second somatosensory area (SII) and cerebellum which are both involved in deliberate control of vocalization. Supporting the model postulated by Wild et. al, the same primary sensory-motor and premotor regions for the control of vocalization are activated in all 3 situations (T, I, and L) which indicates that spontaneous laughter elicitation is also under the control of voluntary motor cortical pathway. Taken together, both groups suggest that motor cortical pathway is involved in generation of both voluntary and involuntary laughter (such as being tickled and even in inhibition of laughter).
Neural Circuitry of Laughter in Chimpanzees
Relatively few studies have been conducted on the higher order frontal lobe control of laughter-like behavior in chimpanzees; however, the neural circuitry that elicits laughter motor excitation remains homologous to a certain degree in both humans and chimpanzees. T. Brown (1915) was the first author to publish his findings on electrical stimulation of different brain areas in chimpanzees. His study involved applying uni-polar electrical current to brain areas of a decerebrated chimpanzee to elicit motor responses. Strikingly, weak electrical stimulation on an area of grey matter around the aqueduct of third ventricle elicited steady respiratory movements at first, but soon changed to a fast and shallow breathing that resembled the laughter-like "pant-pant" sound of chimpanzees. Additionally, the abdominal muscles of both sides of the body appeared to actively contract and relax during the reaction.
Electrical stimulation of ventral part of optic thalamus gave a characteristic alteration of respiration. The breathing becomes vigorous with abdominal muscles taking part in the act. Stimulation of dorsal part of the area, especially to the focal area within thalamas, gave a slowing of respiration, resembling the laughter-like sound vocalization. For both laughter-like responses elicited around third ventricle and optic thalamas, responses outlasted the stimulus that evoked them.
Although there is relatively few scientific study on neural activity of common chimpanzees during laughter, which limits the scope of our website on the subject, based on T. Brown's electrical stimulation study of chimpanzee brain strucutres, the neural pathway for eliciting laughter-like behavior in chimpanzees may share more homologies with humans than one may expect.
Conclusion
With a basic understanding of the neural circuitry invovled in production of laughter, we may compare the physiologic and social aspects of laughter in humans and chimpanzees. Some argue that laughter is the sound of play, whether it be the primal chimpanzee ‘‘pant- pant’’ or the derivative human ‘‘ha-ha’’ (Provine, 2000). Chimpanzee laughter vocalization differs from that of humans: laughing humans chop an outward breath into a series of short (1/15 s), vocal blasts (‘‘ha-ha,’’ etc.) that repeat about every 1/5 s (Provine & Yong, 1991). In contrast, laughing chimpanzees do not parse their exhalations, but produce one laugh sound per outward and inward breath (Provine, 1996). The difference in the vocal frequency output may point to different neural excitation patterns in human and chimpanzee laughter, along with fundamental structural differences in vocalization.
It is noteworthy that chimpanzee laughter occurs almost exclusively during physical contact, or during the threat of such contact, during chasing games, wrestling or tickling (The individual being chased laughs the most). Although people laugh when tickled, most adult human laughter occurs during conversation, typically in the absence of physical contact. The difference in social contexts that elicit human and chimpanzee laughter may be related to the adaptive value of laughter in the two species who face drastically different circumstances of social structure and survival threat.