Bivalve Behaviors
Biology 342: Animal Behavior
Fall 2012
Melissa Ashbaugh, Robin Byron, & Wyatt Gormley
| Home | Escape Behavior | Reproduction and Development | Feeding Behaviors | References | Course Home |
Evolutionary History of Bivalvia
From both a neuroendocrinological and neuroethological perspective, Bivalvia feeding behavior is lacking in complexity compared to most other animal classes. What makes Bivalvia interesting in this regard is the diversity of strategies, ranging from chemoautotrophy in Lucinidae, to carnivory in Poromya. Here, an overview of feeding strategies is offered prior to an extended discussion focusing on carnivory in the genus Poromya.
Bivalvia Feeding Strategies
Below is a summary of known bivalve feeding strategies, listed from least common to most common strategy.
-
Endosymbiosis: Entovalva nhatrangensis, the only known bivalve endosymbiote, is symbiotic with the brown sandfish sea cucumber, Holothuria spinifera. The nhatrangensis lives in the sea cucumber's esophagus, where it consumes diatoms through filter feeding. This species is restricted to shallow waters of Nha Trang Bay, Vietnam, where its host species lives(Bristol et al. 2010).
-
Chemoautotrophy: Several superfamilies, including Solemyidae and Lucinidae, are capable of carbon fixation through symbiosis with sulfur-oxidizing bacteria. These species typically live near deep sea vents or shallow sulfide-rich sediment.(Olu-LeRoy et al. 2004) Bivalves which make use of this strategy tend to have digestive systems which are underdeveloped or non-existent.
-
Carnivorous behavior: Certain bivalves, such as Poromya granulate, have an enlarged siphon, controlled by strong retractor muscles, which allows them to bring food closer to the mouth. This species has fifteen sensory tentacles which emerge from the mantle of the bivalve and grab food. Poromya granulate are found in the subarctic and boreal regions on both sides of the North Atlantic Ocean.
-
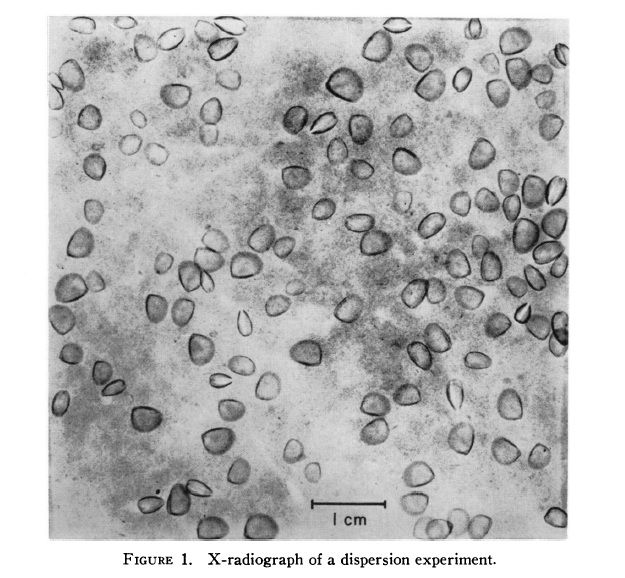
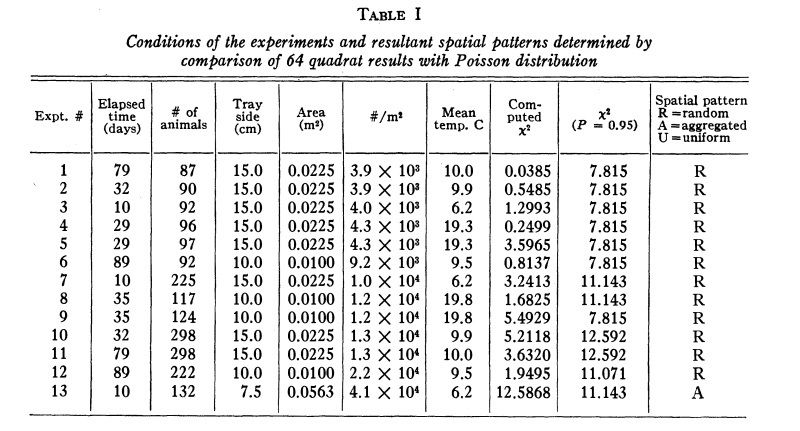
Scavenging: Protobranchia feed by scraping food off of the seabed. This is thought to be the original bivalve feeding strategy before the evolution of specialized gills. Protobranchia typically lack a byssus gland on their foot, although in some occasions they have a nonhomologous byssus gland. One study averaged the distance between Protobranchia individuals and their nearest neighbor using x-radiography.(Figure 1) This measurement was compared to the average distance between a randomly selected point and the nearest Protobranchia specimen to determine that the Protobranchia from their sample were randomly distributed, showing no signs of gregariousness or territoriality.(Table 1) This suggests that selective pressures have not produced nonrandom scavenging in Protobranchia.(Levinton 1972)
-
Filter-Feeding: Perhaps the most common feeding strategy among bivalves is the use of ctenidia to filter phytoplankton from water or from nonnutritive sediment. Filter-feeding bivalves make use of a byssus gland located on their foot to anchor them to the seabed. Filter feeding is the simplest bivalve feeding strategy which constitues recognizable behavior as it involves coordinated movement of cilia controlled by neural pathways.
Carnivorous Poromya
Anatomy of Protobranchia
Mechanism:
In all Poromya species examined, an inhalant siphon is used to get food. Hydraulic pressure changes within the mantle cavity controls extention and enlargment of the of the siphon. Blood is moved from a reservoir into the mantle cavity to produce this pressure change. The siphon is returned to the body by pallial retractor muscles; excess water is transferred to a smaller exhalant siphon. Originally it was thought that the tentacles of Poromya, generally 15 in number, were responsible for prey capture. These have since been observed not to be sticky, as was previously thought, but instead are cilia, mechanosensory organs which allow Poromya to sense the movement of prey.(Morton 1987) There is little data on the exact neural networks that coordinate this behavior, so external motion necessary to cue siphon-based feeding is not well understood.
Ontogeny
The development of carnivorous feeding is not well understood. Few studies characterize the chemosensory cilia that alert Poromya of nearby food, let alone how these cilia affect feeding practices over time. It is not clear at what point Poromya become carnivorous, or whether their diet changes as a result of previous learning.
Phylogeny
Given similar prey capture mechanisms, including the use of an enlarged siphon and ciliary-sense organs, this behavior is likely monophyletic, and produced as early as the Palaeozoic period (540-250 million years ago). As noted, mass extinction then, as well as in the Mesozoic period (250-66 million years ago) make further characterization difficult.
One particular study, conducted by Brian Morton in 1987, undertook a broad phylogenetic discussion of Parilimyidae, Verticordiidae, Poromyidae, and Cuspidariidae, all of which exhibit carnivorous behavior and all of which are found within the taxonomic order Anomalodesmatans. Poromyidae and Cuspidariidae are extant, while Parilimyidae and Verticordiidae are not. The study concludes:
"On the basis of this study, a better picture of phylogenetic affinities in these related bivalves is possible. The stem group seems to be represented by verticordiids and parilimyids. Most verticordiids are of relatively simple plan and their siphonal prey-capture mechanism comparatively unspecialized. Parilimyids and Lyonsiella, however, have a pair of taenioid muscles, enhancing the efficiency of siphonal retraction. Lyonsiella formosa (at least) feeds like poromyids, suggesting a phylogenetic link; the latter, however, being the more morphologically specialized. Cuspidariids, like parilimyids, feed on swimming prey, the long inhalant siphon being everted upwards. This also suggests a phylogenetic link, the cuspidariids being the more specialized members of this group. The Palaeozoic and Mesozoic extinctions of large numbers of anomalodesmatans have left few living descendants and these show widely divergent character traits, making the construction of lineages difficult."(Morton 1987)
Adaptive Value
As discussed in the previous section on phylogeny, carnivory in bivalves has a long history tracing back to the Mesozoic period, possibly further, and is well adapted for extant species.One study examining the stomach contents of Poromya found a partially digested ostracod (the species was not identified). Another specimen contained a relatively intact cirolanid isopod (also unidentified).(Leal 2008)
Scavenging, filter-feeding, and carnivory have existed within Bivalvia since the Mesozoic period, 250 million years ago, demonstrating these behavioral strategies to be of excellent adaptive value. Many questions relating to the origin of these different feeding behavioral practices remain. Of particular note to this discussion is whether one feeding strategy is or was in the past of greater adaptive benefit than other existing strategies. Carnivory is certainly a more efficient feeding strategy than the endosymbiosis found in Entovalva or the chemoautotrophy of Solemyidae and Lucinidae, indicated by Poromya's increased size and larger geographical spread. However, whether it is of greater adaptive value than scavenging or the filter-feeding more common among bivalves is more difficult to determine. Does a greater number of species practicing these strategies suggest greater adaptive value? Does the potential for evolutionary modification between species practicing these strategies speak to the degree of the behaviors' adaptabilities?