Bio 332 - Vascular Plant Diversity:
EXAM 2 KEY
The following answers are given as guidelines to the kinds of information I was looking for. Other correct answers may have been possible for some questions, so you should check with me if you have specific questions.
1. Heterospory has evolved multiple times in the land-plant clade. What advantage(s) does heterospory provide over the ancestral condition? Describe how the advantage(s) of heterospory are (or are not) further modified by the seed plant life cycle.
Homospory results in gametophytes that have both male and female function, and thus intragametophytic self-fertilization is usually possible, with a result that the offspring sporophyte will be completely homozygous. Homosporous gametophytes must balance their allocation to the competing demands of male (sperm) and female (egg) functions. Heterosporous plants produce different spore types that result in single-sex gametophytes, so there is no possibility of intragametophytic selfing, and the spores/gametophytes can evolve towards specialization for their unisexual roles (e.g. the female gametophytes can be produced from larger spores and can themselves grow to a size that better supports the new embryo).
The seed habit is a further elaboration on the differential roles/morphology of the male and female spores/gametophytes, with the megaspore staying resident on the parent sporophyte and the microgametophyte taking on the transport role of traveling to the megapsores’s ovule. It is less obvious how the outcrossing advantage differs between free-sporing and seed heterosporous plants, as both are usually capable of both intergametophytic selfing and outcrossing. Dioecious seed plants do manage to preclude any chance for intergametophytic selfing, but this does not seem to be a general advantage for seed plants, nor a direct consequence of the seed life-cycle.
2. “Since Darwin it has been recognized that the tempo and mode of speciation are greatly influenced by the number and magnitude of genetic changes required for reproductive isolation...". Bradshaw et al (1995; [Nature 376: 762])
Describe an experiment that could be used to estimate the number of genetic changes (= number of loci) for traits that are known to reproductively isolate two species. You should assume the complete genomes are not yet known for the two species. Be sure to specify the data/comparison that would provide the number estimate.
To determine how many genetic loci underlie the evolutionary divergence of traits for two populations/species, you would cross plants from the two groups to make an F1, and then complete further crosses (F2, etc, or back-crosses to the parents) and observe how the traits segregate in the progeny generations. If the trait differences in the original parents are due to a single locus, then both parental phenotypes will be recovered in the F2 at high frequency. If the traits differences are due to many loci, you would not expect to recover the parental types (or you would see them only at low frequency). The higher the number of contributing loci, the lower the trait variance in the F2. An elaboration on this design would be to include genetic markers in the study that identify chromosomal regions, and see how the traits co-vary with particular genetic markers. Each genetic marker that shows a significant association or linkage to variation in a trait marks a "QTL" (quantitative trait locus) for that trait
3. Many botanist have considered the “triploid bridge” to be the more common way for polyploids to form in natural populations. Their view is favored in part by the ecological circumstances of polyploidy formation – when a non-disjunction event occurs during spore formation, the resulting gametophyte (with higher ploidy) will be in the minority in a population made up primarily of normal ploidy gametophytes.
Consider the data shown in the figure to the right, which report the empirically observed pollen cytotypes (chromosome counts for a pollen grain, with ‘X’ = one complement) produced by triploid (sporophyte) pollen parents.
Describe how these data either support or reject
a pathway to polyploid formation that involves
the triploid bridge.
SUPPORT: For the triploid bridge to work, triploid individuals would need to produce spores (and gametophytes) that have not undergone successful reduction from meiosis, so that they can mate with normal, diploid sporophytes and make a tetraploid offspring. Although 3X pollen is of low occurrence (less that 10%), it was observed in the pollen from triploid individuals, suggesting that the triploid pollen (and their sperm) could be combined with a normally reduced egg nucleus in ovules of diploid individuals to produce a tetraploid. If similar failed-meiosis is also occurring in the megapsore production, then normal pollen in the diploid population might combine with unreduced, triploid eggs in triploid sporophytes to make the tetraploid offspring.
REJECT: To complete the triploid bridge, triploid pollen would need to be available. The vast majority (>90%) of pollen being produced by triploid plants is something other than triploid, with an unbalanced (aneuploid) ploidy (3X/2) being the most common form. Thus the likelihood of the minority (<10%) triploid pollen succeeding is minimal.
4. Soltis and Soltis (2007; PNAS 9: 7051-7057) reported that polyploid taxa typically have higher levels of heterozygosity (genetic diversity within loci) than are observed for their diploid parents. Describe how this conclusion relates to i) data they report on the origins of polyploidy species, and ii) how it informs our understanding of the commonness or rarity of polyploid speciation. .
Genetic data involving allo/isozyme electrophoresis and DNA-based markers have shown that the polyploid populations have levels of genetic variation that are inconsistent with a single point of origin from a single hybridization and/or polyploidy event. These data reflect variation within loci and beyond the “fixed heterozygosity” between loci that may arise in allopolyploids when the parent taxa contribute divergent loci. The gene diversity data from non-nuclear markers (cpDNA and mtDNA) are especially convincing, as their uniparental mode of inheritance clearly shows that different parent diploid species served as a seed (=maternal) parent in the polyploidy events.
Diversity within loci (or within the cpDNA and mtDNA genomes) was not expected if polyploidy is a rare event and creates a genetic bottleneck, with only a few individuals contributing their alleles during the formation of the polyploids. Rather, the genetic data show that polyploids have been constituted from the start with lots of genetic variation, in large part because of multiple events creating the polyploid individuals that form the new species. This means the polyploidy event must not be as rare as first envisioned, and it also means the polyploid taxa thus formed might be considered as polyphyletic taxa
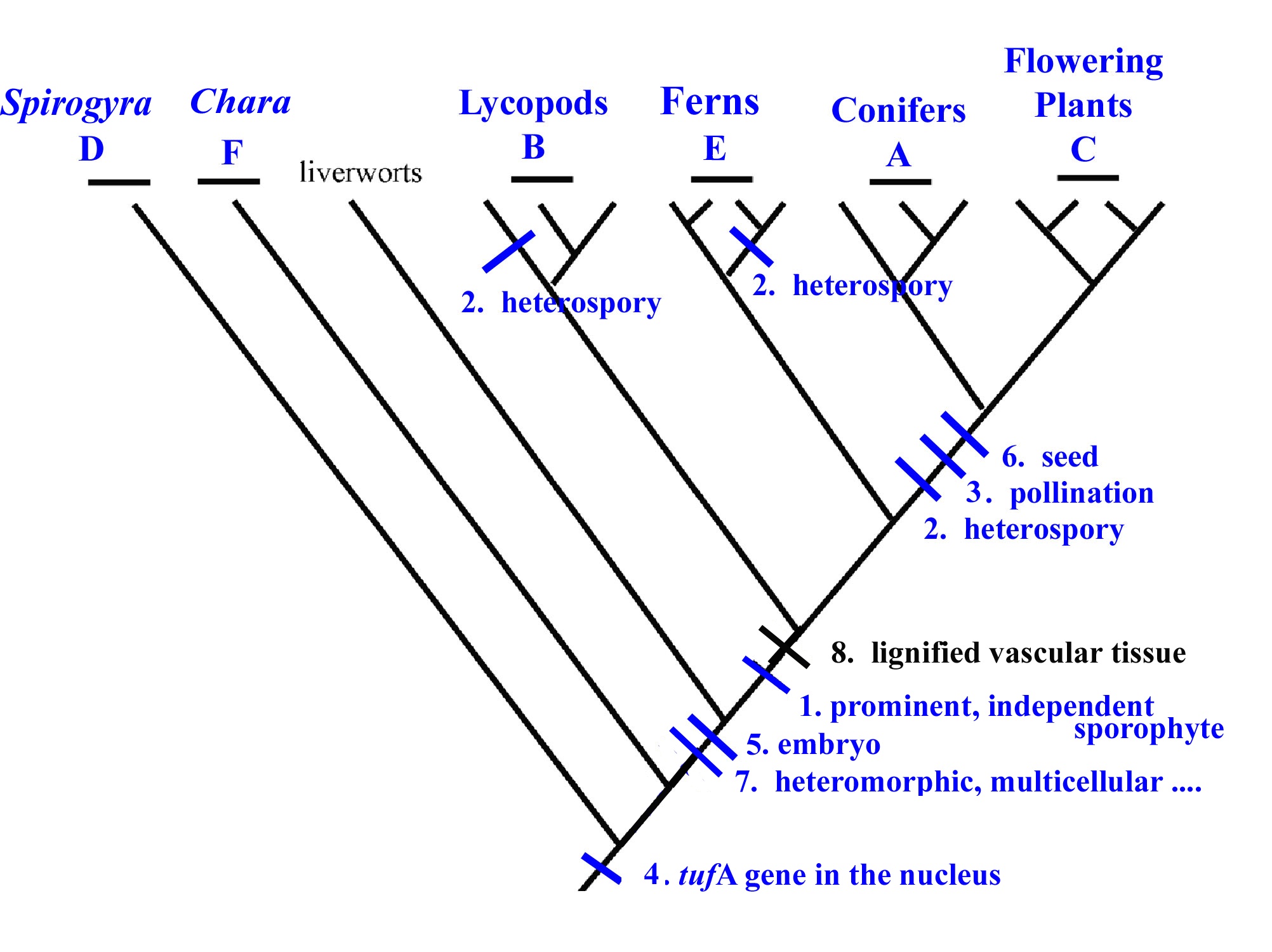
5. On the phylogeny below, use the letter designations (or name) to place all six taxa from the list provided into their appropriate place among the terminal taxa (use as your guide the most recent phylogenetic analyses we discussed in class). Once taxa are assigned, place the gain of each numbered trait onto the phylogeny by putting a hatched mark along the branch(es) during which the trait was gained and assigning the corresponding number to designate that trait (as shown for the gain of the trait 8, lignified vascular tissue). Homoplastic traits should be placed so as to indicate the multiple origins of the trait.
A. Conifers; B. Lycopods; C. Flowering Plants; D. Spirogyra; E. Ferns; F. Chara
1. prominent, independent sporophyte;
2. heterospory;
3. pollination;
4. tuf A chloroplast gene in the nucleus;
5. embryo;
6. seed;
7. heteromorphic, multicellular alternation of generations;
8. lignified vascular tissue;
6. In the case of Mimulus cardinalis (hummingbird pollinated) and M. lewisii (bee-pollinated), the differences between the species include flower color, flower morphology, and the quality and quantity of the nectar reward. Differences between the two species in petal carotenoids were found to result almost entirely from substitution at a single gene locus (yup), with the M. lewisii allele dominant to the M. cardinalis allele. Schemske and Bradshaw (1999; PNAS 96: 11910-11915) observed the following data when watching pollinators visit F1 and F2 plants in the field (L and C refer to the M. lewisii and M. cardinalis alleles, respectively, as determined by the genetic markers linked to the yup QTL).
Describe how these results from the pollinator observations combine with the genetic results and contribute to an understanding of the tempo and mode (geographic) of speciation in this case.
Tempo in this case refers to the speed of the speciation process, while mode refers to the geographic circumstances of the populations that are speciating. The data on pollinator preferences show that for a trait that is affected primarily by a single locus, the phenotypic differences can have a very pronounced consequence for pollinator visitation rates and therefore for pre-mating reproductive isolation. A major gene locus that provides reproductive isolation could provide for a very rapid evolutionary divergence leading to speciation. Genes of major effect that lead to rapid divergence are an important element of the sympatric geographic mode, so these data could support a model of sympatric speciation. Rapid changes of this sort could also arise in allopatric populations, however, so the allopatric mode of speciation is not precluded. These data show primarily an effect of yup on insect visitation, so the single locus is unlikely, on its own, to eliminate visitation by hummingbirds to all genotypes. Reproductive isolation for sympatric populations might therefore require additional genetic changes to lead to hummingbirds limiting visitation to the “M. lewisii” plants with a yup L allele, so the sympatric explanation is probably less likely than an allopatric scenario based on these data for the yup locus
7. Describe how the correct identification of the algal ancestor to the land plants has resolved conflicting ideas about the evolutionary origin of the type of alternation of generations that is observed for all land plants. In your explanation, be sure you account for the unique feature(s) (synapomorphy(ies)) that define(s) the plant kingdom as distinct from its algal relatives
Competing ideas were advanced about either a diplobiontic algal ancestor (so plants inherited from algae an alternation of generations with multicelllular diploid and haploid phases), or a haplontic algal species (with no multicellular diploid stage). The first hypothesis was considered to include the possible independent origin from different algae of vascular plants and non-vascular plants. The second hypothesis has been supported by a great many shared, derived characters of land plants and algae, with Charophycean algae as the closest relatives of land plants. These algae have a diploid zygote that undergoes meiosis without first dividing mitotically, and they lack a multicellular diploid stage. Given this ancestor, land plants evolved an alternation of generations in which they added mitotic divisions and delayed meiosis, introducing a multicellular diploid stage (the sporophyte) which had the capacity for diploid gene expression and which could produce more meiotic spores. The embryo and the heteromorphic diplobiontic life cycle are the defining characteristics for the plant kingdom. The zygote for some Charophycean algae is retained by the parent gametophyte. If the zygote that evolved the new mitotic divisions was also retained on the egg-parent gametophyte, this could explain the evolution of the embryo stage of the land plant life cycle. Later diversification of the land plants increased the importance of this new sporophyte stage of the life cycle.
8. Both Niklas et al. (1985) and Steeves (1983) commented on the remarkable length of time between the first appearance of seeds in the fossil record and the ascendancy of seed plants to become the dominant group of the terrestrial flora. Steeves suggested a parallel with vertebrate evolution, where the reptiles appeared in the Carboniferous, but did no come to dominate until after the Permian and throughout the Mesozoic, despite “superior adaptations for terrestrial reproduction” in the reptiles.
Describe the superior adaptations for terrestrial reproduction provided by the seed habit, and evaluate two of the hypotheses offered by Niklas et al. and/or Steeves to account for the apparent slow rise to dominance shown by seed plants.
Seed plants differ from non-seed pants in retaining their spores, dispersing juvenile diploid sporophytes (rather than spores), and not relying on free-swimming sperm in the external environment to provide the gamete transport leading to fertilization (they use the pollen grain to get the sperm to the vicinity of the egg in the ovule). The freedom from the requirement for external water to allow sperm movement through the environment allows these plants to reproduce in areas where free water in the environment is scarce. The dispersal of a multicellular, diploid sporophyte (rather than a haploid spore) provides for the establishment of new plants with a propagule that has diploid gene expression and a “head start” in the form of multicellular tissues and nutrition. While the spore also provides nutrition for the start of the gametophyte (and can also provide for dormancy), seeds tend to be much larger and are often dispersed with more provisions.
Explanations for the slow rise include i) the slow acquisition of morphological/physiological adaptations that could allow for growth in drier habitats, ii) the biases present in the fossil record due to which habitats are more often preserved (tends to be low elevation, wet habitats where sediments are deposited), and/or iii) a distinction between species diversity (Niklas et al. data) and ecological prominence. Early seed plants may have existed in dense stands over wide areas in ways that provided outcrossing and high gene flow, limiting diversification rates.
9. Allozyme variation detected through protein electrophoresis has provided researchers with the ability to estimate the intragametophytic selfing rate for natural populations of free-sporing plants.
i) describe how the electrophoretic data are used to estimate the intragametophytic selfing rate,
ii) indicate any problems with this method that might cause it to provide a biased estimate of the actual rate of intragametophytic selfing, and
iii) account for the observation that some populations are observed where the sampled sporophytes are all of a single genotype, yet the intragametophytic selfing rate estimate is zero (i.e., what kind of reproduction has produced the sampled sporophytes?)
i) Intragametophytic selfing results in zygotes (=sporophytes) that are completely homozygous at all loci. Allozyme genetic markers are co-dominant, and heterozygotes can be distinguished from both homozygotes. By genotyping sporophytes at many allozyme loci, we can determine which individuals are heterozygous for at least one locus (and thus, they could not have been a zygote produced by intragametophytic selfing). The frequency of individuals homozygous at all surveyed loci is then an estimate of the intragametophytic selfing rate.
ii) some of these homozygous sporophytes are likely to have been produced by matings between gametophytes (either outcrosssing or intergametophytic selfing) that had the same allele at all surveyed loci. They are completely homozygous at the sampled loci, but they were not produced by intragametophytic selfing (other loci which were not sampled would show this to be the case). Thus, this estimate is a biased, over-estimate of the actual rate of intragametophytic selfing.
iii) Because the intragametophytic selfing rate is estimated to be zero, the single observed genotype must be heterozygous at one or more of the sampled loci. The likelihood of sampling only heterozygotes in a population that is reproducing sexually is extremely small (homozygotes for both alleles should be observed), so this either indicates an evolutionary force like natural selection is also affecting this locus (perhaps only the heterozygotes can survive) or the sampled plants have been produced not through sexual reproduction, but rather through asexual reproduction (clonal spread of a single, heterozygous genotype)