Fish Schooling
Biology 342 Fall 08
Ontogeny
If fish have the basic locomotor and sensory capabilities necessary for schooling behavior at hatching, what necessitates the development of this behavior?
Development of schooling behavior has been observed in many species concurrently with metamorphosis into juvenile form and schooling continues to be refined into adulthood (Gallego and Heath 1994, Nakayama 2007). However, a critical mechanism for schooling, the central nervous system, is not fully developed in the larval stage, and early and imperfect schooling poses potentially greater risk than not schooling at all. Additionally, in a study by Nakayama, schooling behavior occurred a full two weeks earlier than some observed benefits of schooling, such as social transmission conveyed by visual cues, suggesting that another factor must be driving the development of this behavior.
So why risk it?
Metamorphosis brings different habitat and prey preferences, which influence the relative value and detrimental effects of schooling. Physiological changes associated with metamorphosis, such as increased pigmentation which decreases opacity (see larval and adult forms, below) and swimming speed, alter the conspicuousness of a lone fish (Langsdale 1993) and may make schooling more advantageous.
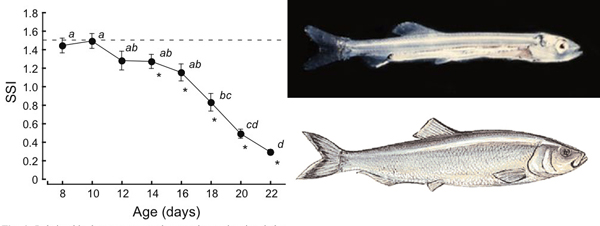
Separation swimming index (SSI) in pairs of fish varies by age, as indicated by this graph from Nakayama. The dotted line indicates the average value of SSI when swimming behavior is totally random (1.49). The SSI begins to steadily decline at 14 days posthatching (dph) and sharply after 16 dph, at which point the fish had completed metamorphosis from the larval stage (top) to the adult form (bottom).
Is schooling learned?
Fish can make subtle and adaptive adjustments to their antipredator repertoire,
and that learning, both individual and social, is an important survival tool for fish.
- Jennifer L. Kelley and Anne E. Magurran
Although there is clearly a huge selective pressure on fish species to develop predator-eluding behaviors, a great deal of evidence points to a learned aspect of these behaviors as well. Much schooling behavior can be modified with experience from larval stages into adulthood.
Other aspects of schooling, such as choosing the right group to join, are mediated by learning as well. When engaged in with kin, shoaling, a less formal aggregation behavior than schooling, has been shown to lead to less aggressive behaviors, more altruistic behaviors, and more stable dominance hierarchies. A study by Frommen et al., showed that adult sticklebacks prefer to shoal with kin and familiars. When kin and non-kin are reared together, the ability to recognize non-kin is diminished, indicating a learned basis for kin-recognition.
Or innate?
Evidence for the innateness of schooling behavior includes the variability in degree of schooling depending on a given populations risk factors. Guppies, sticklebacks, and minnows from high-predation environments school significantly more than low-predation area fish, even after several generations of laboratory rearing. High-risk populations also show greater phenotypic plasticity in learning these behaviors. (Kelley and Magurran 2003)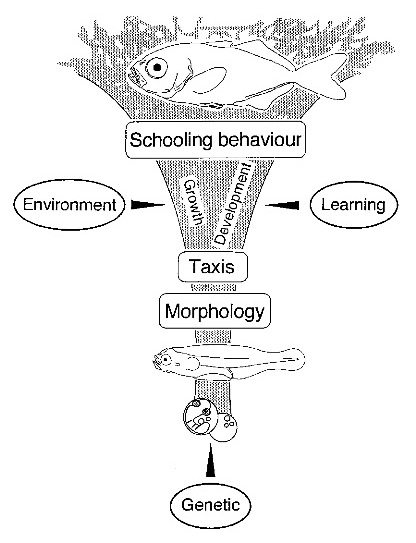
Environmental factors are at work, too.
No matter whether a schooling is innate or learned, environmental factors strongly influence the development of this behavior. Both pre- and post-hatching manipulation of the fishs interior and exterior environment were found to have deleterious effects on schooling.
An experiment by Williams and Coutant showed that sublethal exposure of smelt eggs and larva to heat, simulating thermal plumes, severely retarded and modified schooling behavior. Not only was locomotor ability diminished, but schools were less uniform, formed later, and dispersed more quickly.
A post-hatching manipulation, the dietary depletion of decosahexaenoic acid (DHA), which is critical to the development of the central nervous system, completely wiped out schooling behavior in adult carangid fish. (Masuda and Tsukamoto 1999)
Masuda and Tsukamoto 1999