Laughter in Chimpanzees and Humans - Biology 342 Fall 2012
By Yuan Xue, Soso
What Is the Evolutionary Relationship Between Human and Chimpanzee Laughter Behavior?
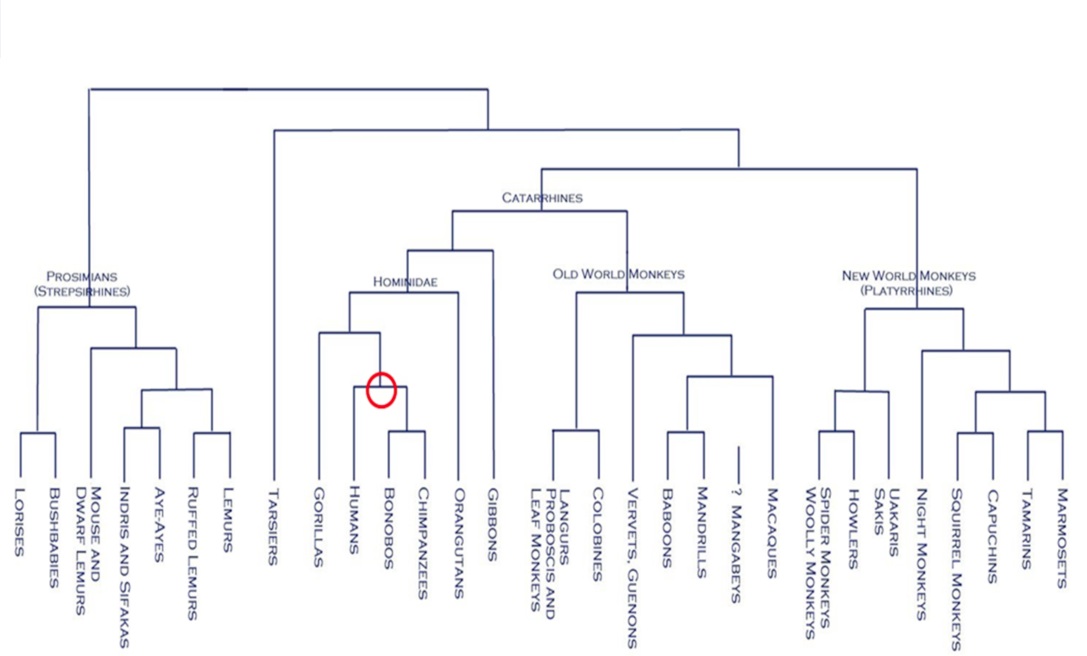 Common Chimpanzees and humans are relatively closely related; our most recent common ancestor dates to be about 6 million years ago. Genetically speaking, the sequence divergence of humans and chimpanzees in DNA is estimated to be about 4.85% (Britten, 2002). Most studies suggested that humans and chimpanzees share many similar social behaviors and patterns.
Common Chimpanzees and humans are relatively closely related; our most recent common ancestor dates to be about 6 million years ago. Genetically speaking, the sequence divergence of humans and chimpanzees in DNA is estimated to be about 4.85% (Britten, 2002). Most studies suggested that humans and chimpanzees share many similar social behaviors and patterns.
In fact, a study on primate laughter behavior revealed strong evidence that laughter, especially when induced by tickling, is homologous between great apes (orangutans, gorillas, chimpanzes, and bonobos) and humans in terms of source of vocalization and social context; it supports the more general postulation of phylogenetic continuity from nonhuman displays to human emotional expressions of laughter-like behavior (Ross et. al, 2009). Distinctively human laughter characteristics such as predominantly regular, stable voicing and consistently egressive airflow are found to be traceable to characteristics of shared ancestors with great apes.
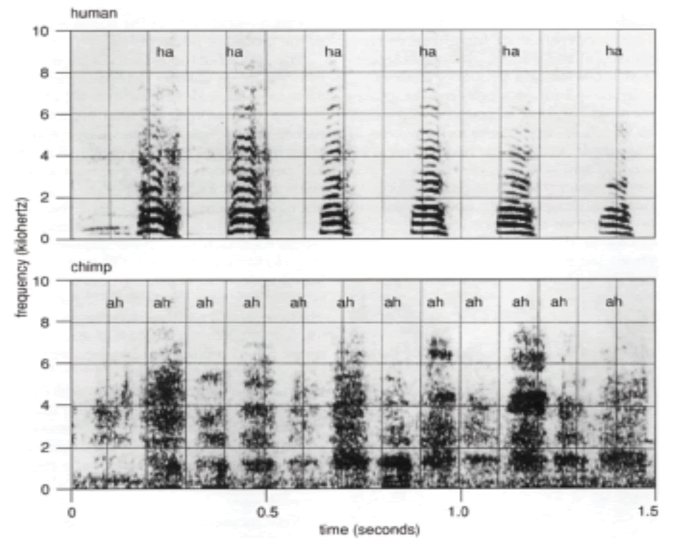
Frequency Spectra of human (top) and chimpanzee (bottom) reveals similarity in voicing production via stable and consistent airflow exchange. Human laughter is, however, distinguished from chimpanzee laughter by its sharply defined onset and offset of the voiced, vowel-like notes.
Despite certain shared critical features, humans and chimpanzees also reveal their fundamental differences when they laugh. One such fundamental difference is the fact that chimpanzee laughter sound lacks discrete vowel and instead produces breathy “pant-pant” sound. Some scientists argue that the differences in laughter can be attributed to the emergence of speech and bipedalism. For most primates, quadrupedalism presents a critical constraint on the evolution of speech (Provine, 1996, 2000). Chimpanzee vocalization is captive to an inflexible neuromuscular system that is closely tied to the ancient and essential labor of breathing. Although not obligate quadrupeds, chimpanzees share constraints typical of other four-legged animals, whose breathing and running are closely synchronized (one stride per breath) in order to brace the thorax for forelimb impacts. Without inflated lungs, the thorax is a floppy, air-filled bag, which is why we humans also hold our breath and may even grunt to push or lift a heavy weight. Respiratory constraints are at least as critical to the emergence of speech as the more traditionally studied structures of the tongue, larynx, and vocal tract. Ultimately, speech requires the cessation of breathing and redirection of the respiratory apparatus in the service of sound making. Being able to do so, allows humans to effectively vocalize speech and laugh by producing more highly defined vowel-like notes than chimpanzees.
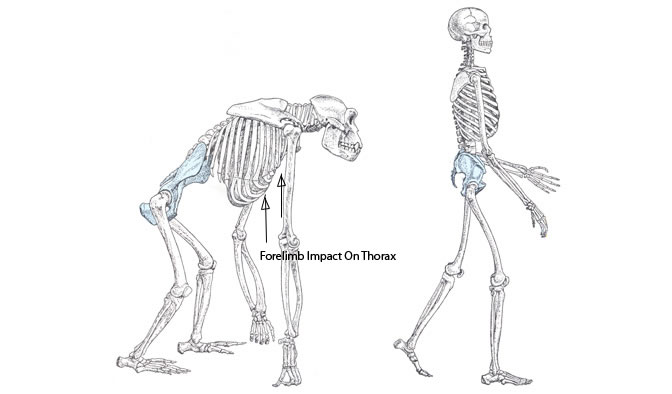 Bipedalism makes the difference
Bipedalism makes the difference
The evolution of bipedalism permitted flexibility in the co- ordination of breathing, running, and vocalizing. This is the basis of the bipedal (‘‘walkie-talkie’’) theory of speech evolution (Provine, 2000). A bipedal human runner, for example, may employ a variety of strides per breath: The ratio can be 4:1, 3:1, 5:2, 2:1, 3:2, or 1:1, with 2:1 being the most common (Bramble & Currier, 1983). Freed of the necessarily rigid 1:1 link between stride and breath characteristic of quadrupeds, humans evolved a vocal system in which individual sounds were no longer tied to single breaths, permitting the subsequent natural selection for speech and, incidentally, our species’ characteristic ‘‘ha-ha’’ laugh.